

Ku recruits the XRCC4-ligase IV complex to DNA ends
- PMID: 10757784
- PMCID: PMC85565
- DOI: 10.1128/MCB.20.9.2996-3003.2000
Ku recruits the XRCC4-ligase IV complex to DNA ends
Abstract
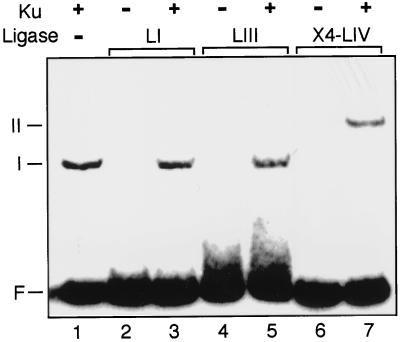
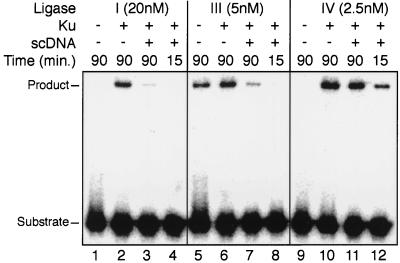
Genetic experiments have determined that Ku, XRCC4, and ligase IV are required for repair of double-strand breaks by the end-joining pathway. The last two factors form a tight complex in cells. However, ligase IV is only one of three known mammalian ligases and is intrinsically the least active in intermolecular ligation; thus, the biochemical basis for requiring this ligase has been unclear. We demonstrate here a direct physical interaction between the XRCC4-ligase IV complex and Ku. This interaction is stimulated once Ku binds to DNA ends. Since XRCC4-ligase IV alone has very low DNA binding activity, Ku is required for effective recruitment of this ligase to DNA ends. We further show that this recruitment is critical for efficient end-joining activity in vitro. Preformation of a complex containing Ku and XRCC4-ligase IV increases the initial ligation rate 20-fold, indicating that recruitment of the ligase is an important limiting step in intermolecular ligation. Recruitment by Ku also allows XRCC4-ligase IV to use Ku's high affinity for DNA ends to rapidly locate and ligate ends in an excess of unbroken DNA, a necessity for end joining in cells. These properties are conferred only on ligase IV, because Ku does not similarly interact with the other mammalian ligases. We have therefore defined cell-free conditions that reflect the genetic requirement for ligase IV in cellular end joining and consequently can explain in molecular terms why this factor is required.
Figures
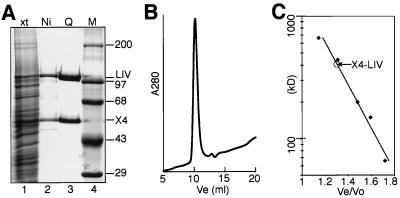
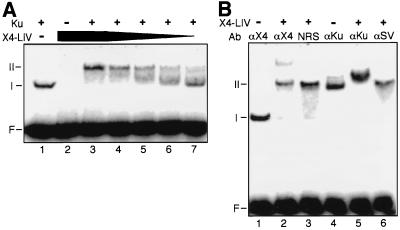
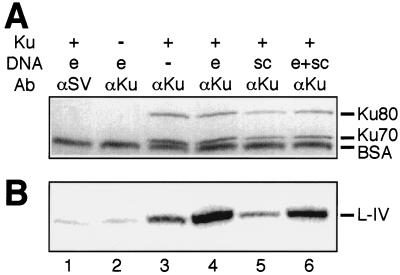
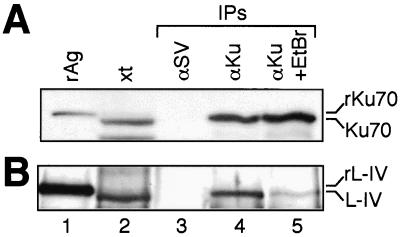

References
-
- Bogue M A, Wang C, Zhu C, Roth D B. V(D)J recombination in Ku86-deficient mice: distinct effects on coding, signal, and hybrid joint formation. Immunity. 1997;7:37–47. - PubMed
-
- Bryans M, Valenzano M C, Stamato T D. Absence of DNA ligase IV protein in XR-1 cells: evidence for stabilization by XRCC4. Mutat Res. 1999;433:53–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous