

Cyclic AMP signaling functions as a bimodal switch in sympathoadrenal cell development in cultured primary neural crest cells
- PMID: 10757785
- PMCID: PMC85567
- DOI: 10.1128/MCB.20.9.3004-3014.2000
Cyclic AMP signaling functions as a bimodal switch in sympathoadrenal cell development in cultured primary neural crest cells
Abstract
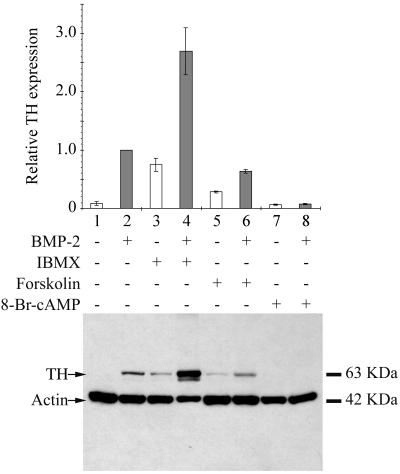
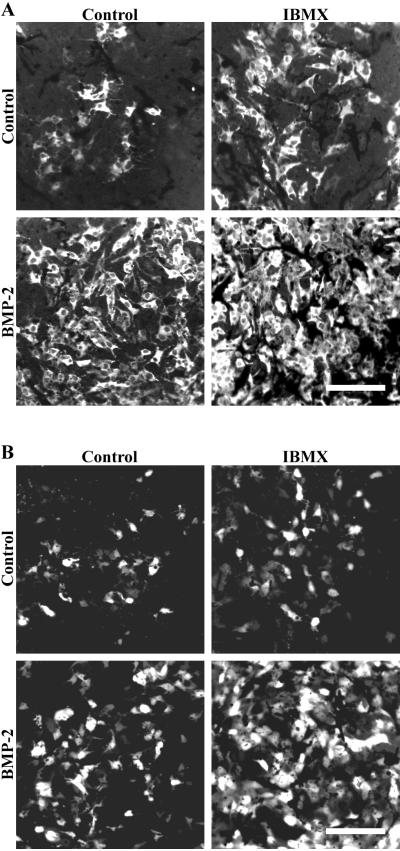
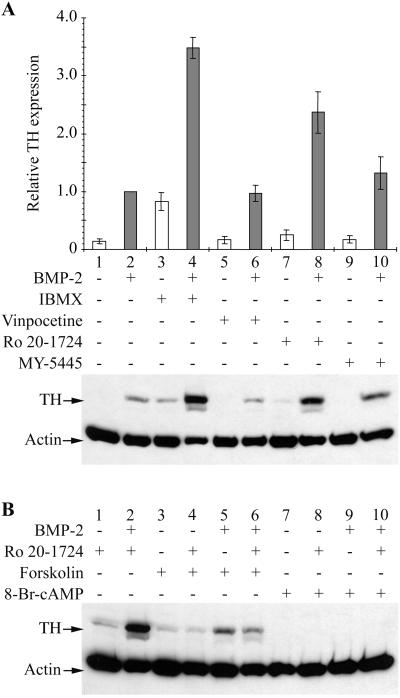
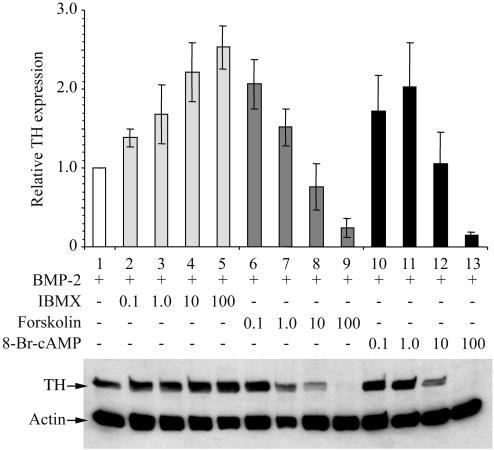
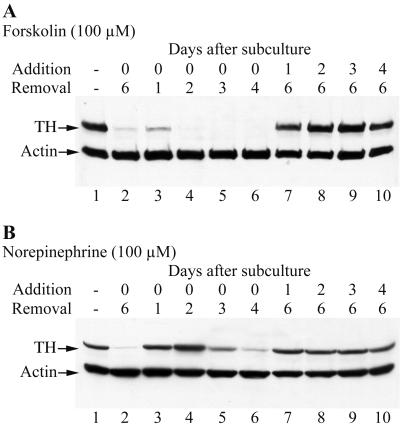
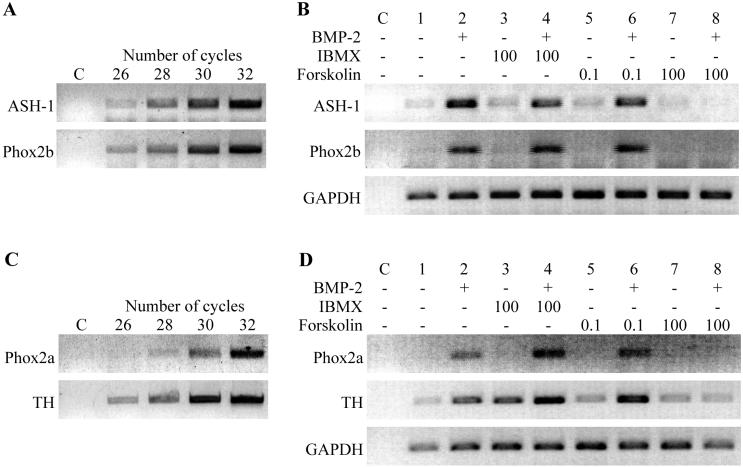
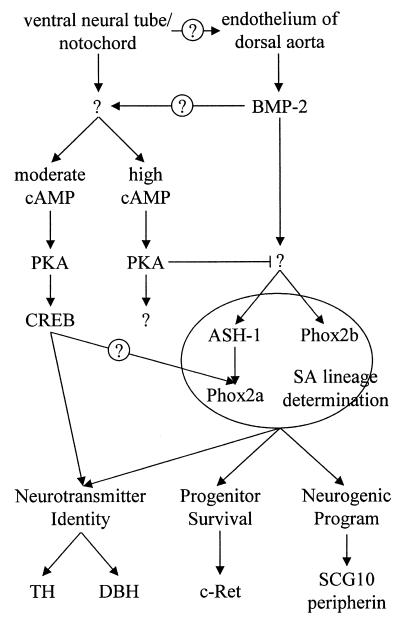
Cells of the vertebrate neural crest (crest cells) are an invaluable model system to address cell fate specification. Crest cells are amenable to tissue culture, and they differentiate to a variety of neuronal and nonneuronal cell types. Earlier studies have determined that bone morphogenetic proteins (BMP-2, -4, and -7) and agents that elevate intracellular cyclic AMP (cAMP) stimulate the development of the sympathoadrenal (SA, adrenergic) lineage in neural crest cultures. To investigate whether interactive mechanisms between signaling pathways influence crest cell differentiation, we characterized the combinatorial effects of BMP-2 and cAMP-elevating agents on the development of quail trunk neural crest cells in primary culture. We report that the cAMP signaling pathway modulates both positive and negative signals influencing the development of SA cells. Specifically, we show that moderate activation of cAMP signaling promotes, in synergy with BMP-2, SA cell development and the expression of the SA lineage-determining gene Phox2a. By contrast, robust activation of cAMP signaling opposes, even in the presence of BMP-2, SA cell development and the expression of the SA lineage-determining ASH-1 and Phox2 genes. We conclude that cAMP signaling acts as a bimodal regulator of SA cell development in neural crest cultures.
Figures
References
-
- Ahn H S, Crim W, Romano M, Sybertz E, Pitts B. Effects of selective inhibitors on cyclic nucleotide phosphodiesterases of rabbit aorta. Biochem Pharmacol. 1989;38:3331–3339. - PubMed
-
- Allan I J, Newgreen D F. Catecholamine accumulation in neural crest cells and the primary sympathetic chain. Am J Anat. 1977;149:413–421. - PubMed
-
- Anderson D J. Cellular and molecular biology of neural crest cell lineage determination. Trends Genet. 1997;13:276–280. - PubMed
-
- Anderson D J. Molecular control of cell fate in the neural crest: the sympathoadrenal lineage. Annu Rev Neurosci. 1993;16:1291–1258. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous