

TAK1 participates in c-Jun N-terminal kinase signaling during Drosophila development
- PMID: 10757786
- PMCID: PMC85571
- DOI: 10.1128/MCB.20.9.3015-3026.2000
TAK1 participates in c-Jun N-terminal kinase signaling during Drosophila development
Abstract
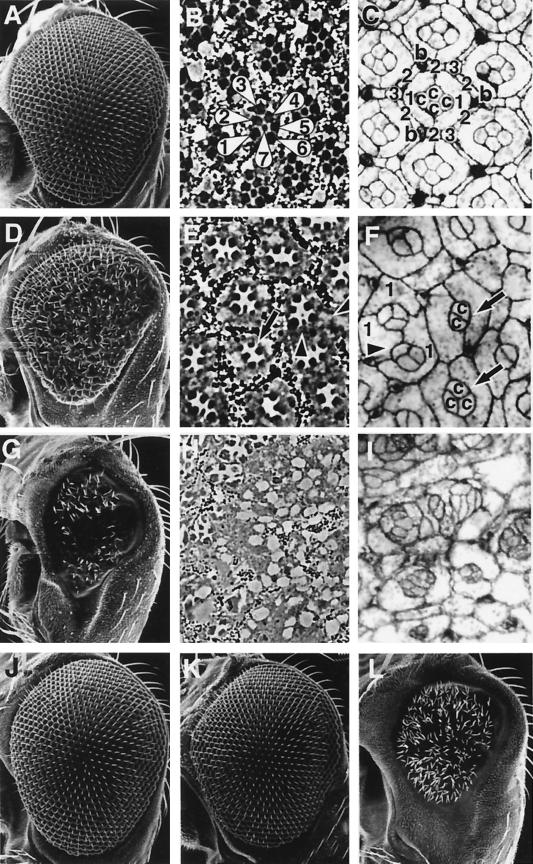
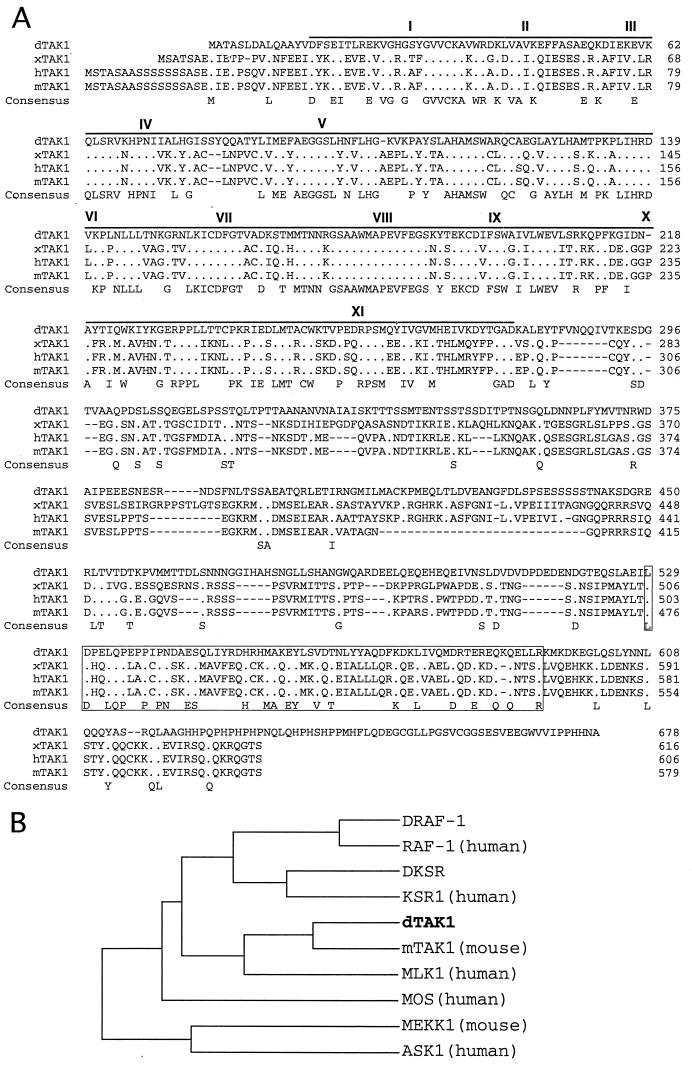
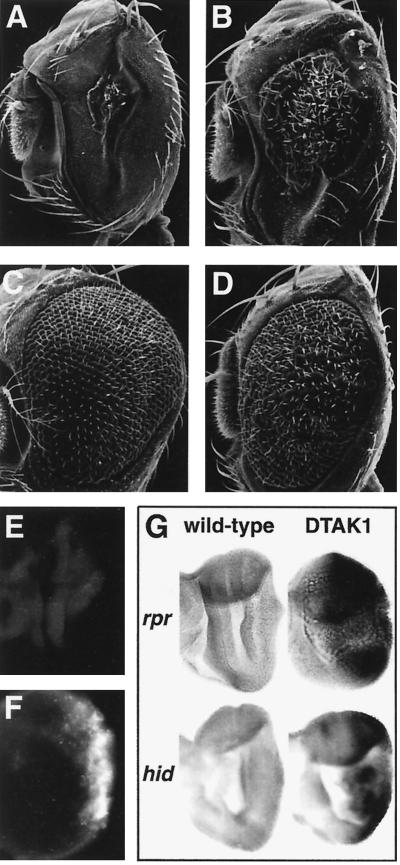
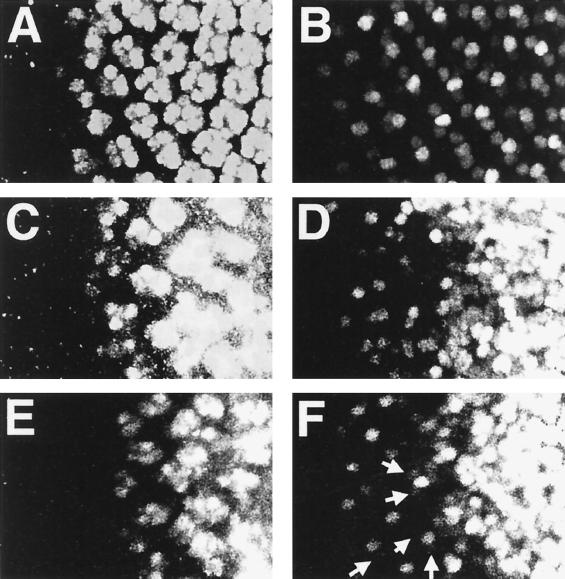
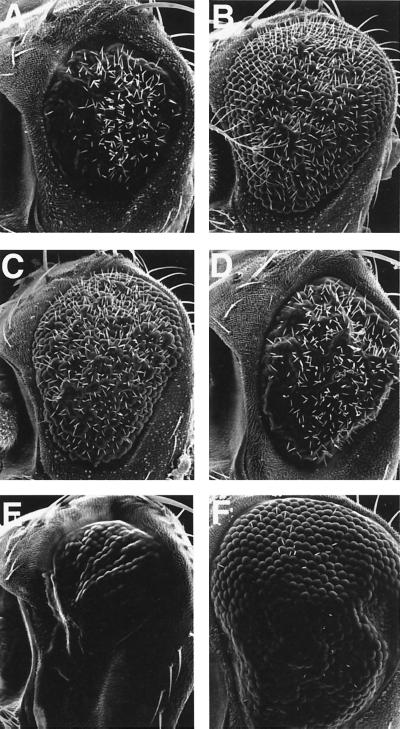
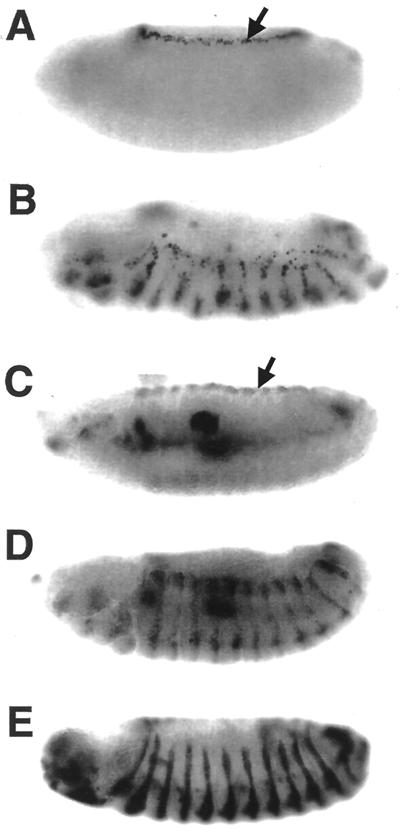
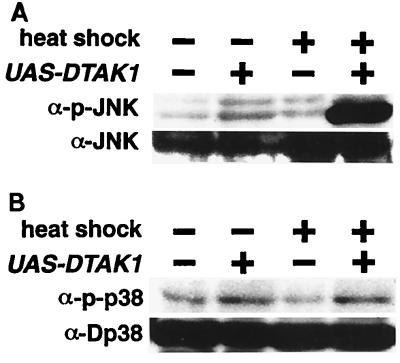
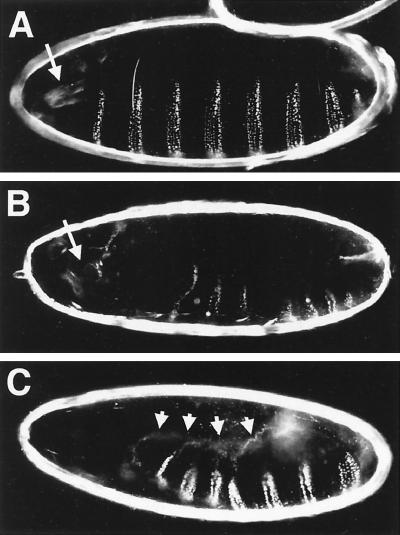
Transforming growth factor beta (TGF-beta)-activated kinase 1 (TAK1) is a member of the MAPKKK superfamily and has been characterized as a component of the TGF-beta/bone morphogenetic protein signaling pathway. TAK1 function has been extensively studied in cultured cells, but its in vivo function is not fully understood. In this study, we isolated a Drosophila homolog of TAK1 (dTAK1) which contains an extensively conserved NH(2)-terminal kinase domain and a partially conserved COOH-terminal domain. To learn about possible endogenous roles of TAK1 during animal development, we generated transgenic flies which express dTAK1 or the mouse TAK1 (mTAK1) gene in the fly visual system. Ectopic activation of TAK1 signaling leads to a small eye phenotype, and genetic analysis reveals that this phenotype is a result of ectopically induced apoptosis. Genetic and biochemical analyses also indicate that the c-Jun amino-terminal kinase (JNK) signaling pathway is specifically activated by TAK1 signaling. Expression of a dominant negative form of dTAK during embryonic development resulted in various embryonic cuticle defects including dorsal open phenotypes. Our results strongly suggest that in Drosophila melanogaster, TAK1 functions as a MAPKKK in the JNK signaling pathway and participates in such diverse roles as control of cell shape and regulation of apoptosis.
Figures
References
-
- Adachi-Yamada T, Nakamura M, Irie K, Tomoyasu Y, Sano Y, Mori E, Goto S, Ueno N, Nishida Y, Matsumoto K. p38 mitogen-activated protein kinase can be involved in transforming growth factor β superfamily signal transduction in Drosophila wing morphogenesis. Mol Cell Biol. 1999;19:2322–2329. - PMC - PubMed
-
- Ashburner M. Drosophila: a laboratory manual. New York, N.Y: Cold Spring Harbor Laboratory Press; 1989.
-
- Boutros M, Paricio N, Strutt D I, Mlodzik M. Dishevelled activates JNK and discriminates between JNK pathways in planar polarity and wingless signaling. Cell. 1998;94:109–118. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous