

A Drosophila ESC-E(Z) protein complex is distinct from other polycomb group complexes and contains covalently modified ESC
- PMID: 10757791
- PMCID: PMC85591
- DOI: 10.1128/MCB.20.9.3069-3078.2000
A Drosophila ESC-E(Z) protein complex is distinct from other polycomb group complexes and contains covalently modified ESC
Abstract
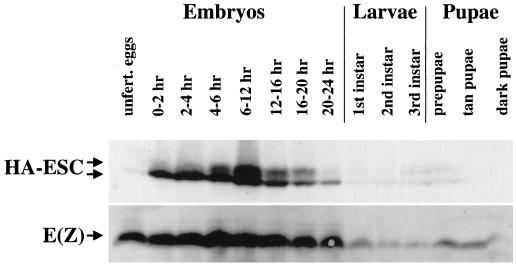
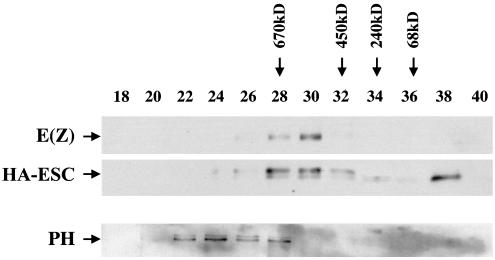
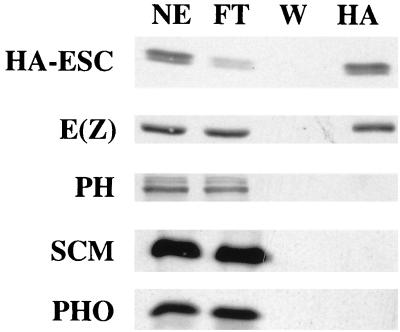
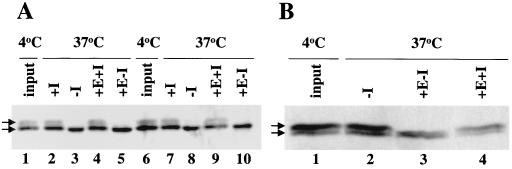
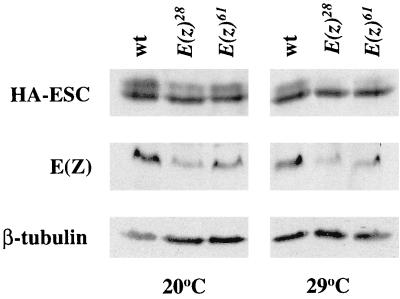
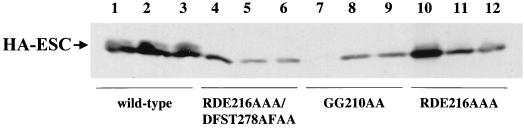
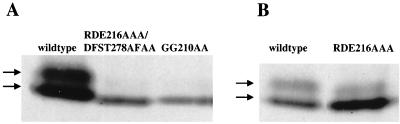
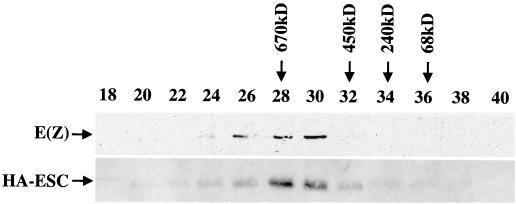
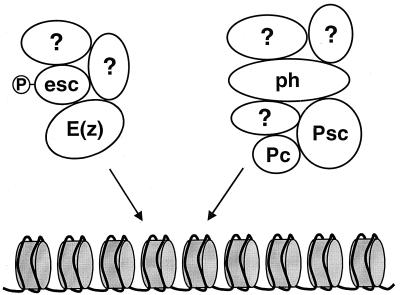
The extra sex combs (ESC) and Enhancer of zeste [E(Z)] proteins, members of the Polycomb group (PcG) of transcriptional repressors, interact directly and are coassociated in fly embryos. We report that these two proteins are components of a 600-kDa complex in embryos. Using gel filtration and affinity chromatography, we show that this complex is biochemically distinct from previously described complexes containing the PcG proteins Polyhomeotic, Polycomb, and Sex comb on midleg. In addition, we present evidence that ESC is phosphorylated in vivo and that this modified ESC is preferentially associated in the complex with E(Z). Modified ESC accumulates between 2 and 6 h of embryogenesis, which is the developmental time when esc function is first required. We find that mutations in E(z) reduce the ratio of modified to unmodified ESC in vivo. We have also generated germ line transformants that express ESC proteins bearing site-directed mutations that disrupt ESC-E(Z) binding in vitro. These mutant ESC proteins fail to provide esc function, show reduced levels of modification in vivo, and are still assembled into complexes. Taken together, these results suggest that ESC phosphorylation normally occurs after assembly into ESC-E(Z) complexes and that it contributes to the function or regulation of these complexes. We discuss how biochemically separable ESC-E(Z) and PC-PH complexes might work together to provide PcG repression.
Figures
References
-
- Alkema M J, Bronk M, Verhoeven E, Otte A, van't Veer L J, Berns A, van Lohuizen M. Identification of Bmi1-interacting proteins as constituents of a multimeric mammalian Polycomb complex. Genes Dev. 1997;11:226–240. - PubMed
-
- Bornemann D, Miller E, Simon J. The Drosophila Polycomb group gene Sex comb on midleg (Scm) encodes a zinc finger protein with similarity to polyhomeotic protein. Development. 1996;122:1621–1630. - PubMed
-
- Brown J L, Mucci D, Whiteley M, Dirksen M L, Kassis J A. The Drosophila Polycomb group gene pleiohomeotic encodes a DNA binding protein with homology to the transcription factor YY1. Mol Cell. 1998;1:1057–1064. - PubMed
-
- Brunk B P, Martin E C, Adler P N. Drosophila genes Posterior sex combs and suppressor two of zeste encode proteins with homology to the murine bmi-1 oncogene. Nature. 1991;353:351–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases