

Cid1, a fission yeast protein required for S-M checkpoint control when DNA polymerase delta or epsilon is inactivated
- PMID: 10757807
- PMCID: PMC85617
- DOI: 10.1128/MCB.20.9.3234-3244.2000
Cid1, a fission yeast protein required for S-M checkpoint control when DNA polymerase delta or epsilon is inactivated
Abstract
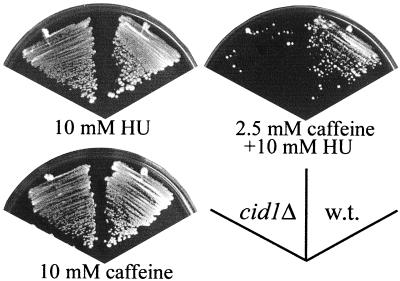
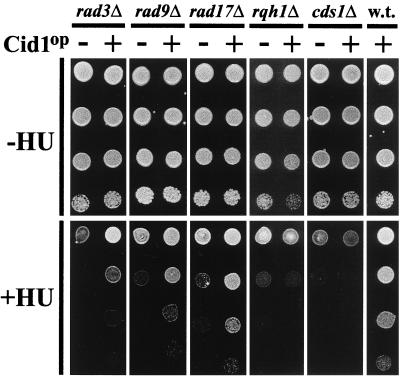
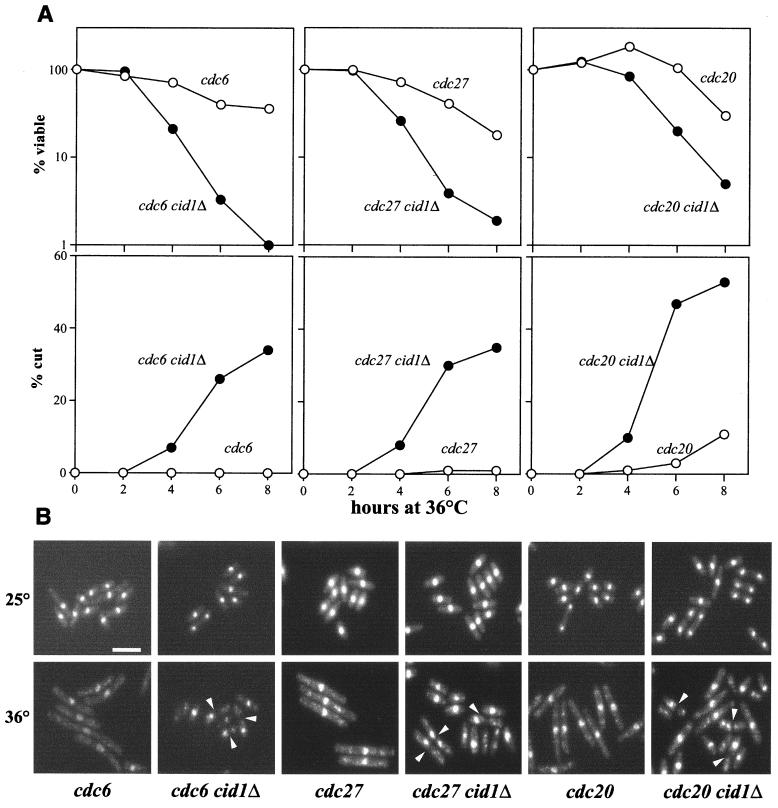
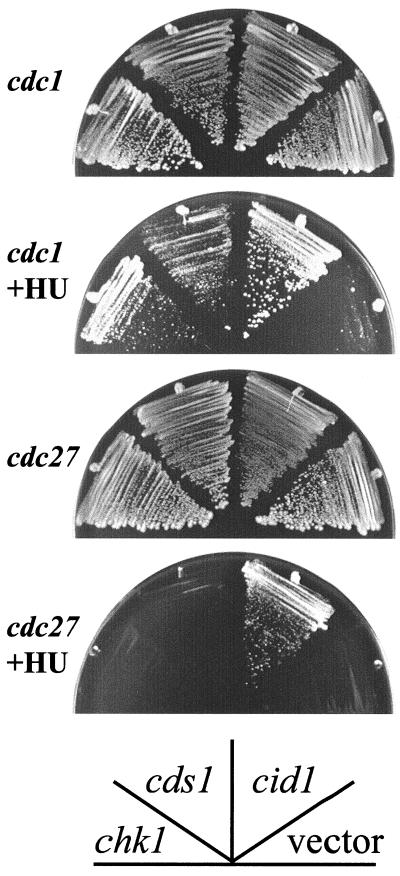
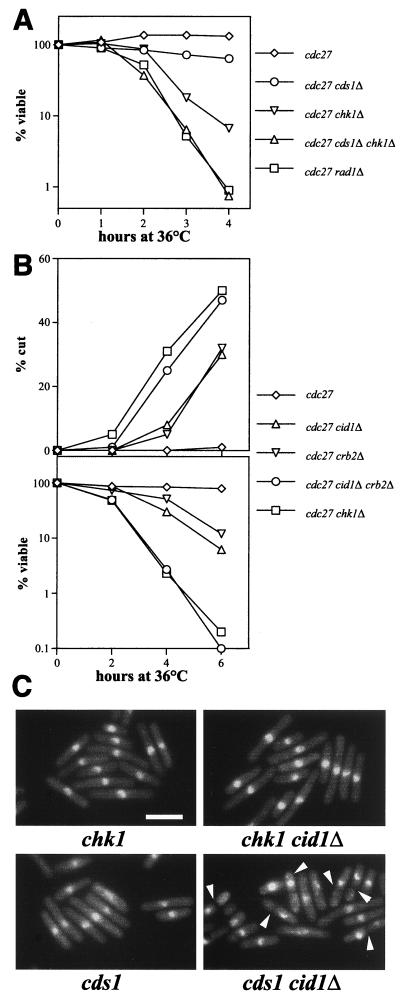
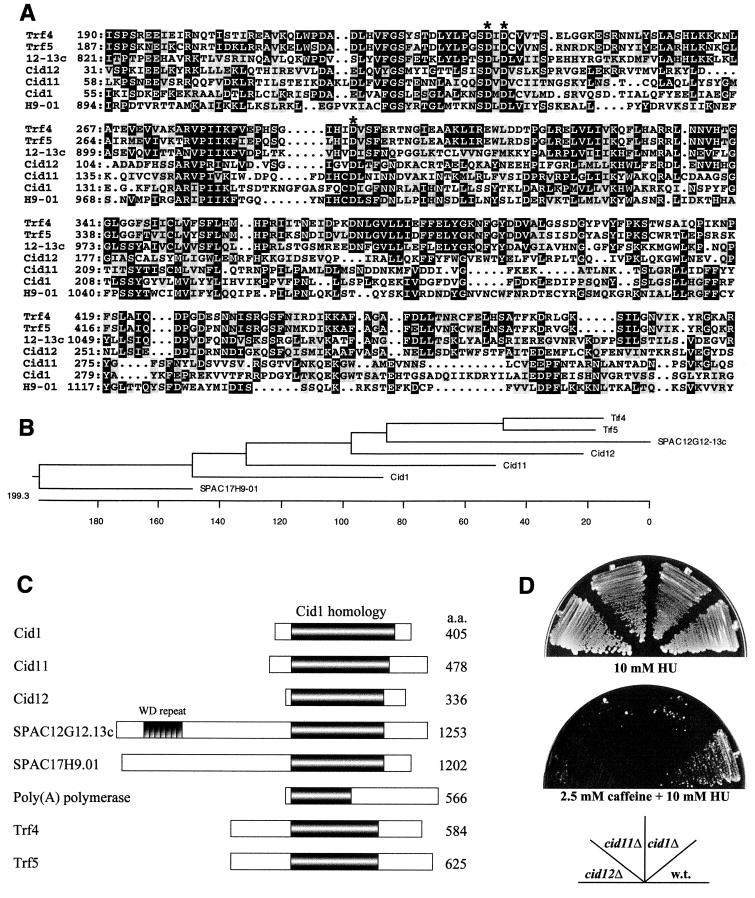
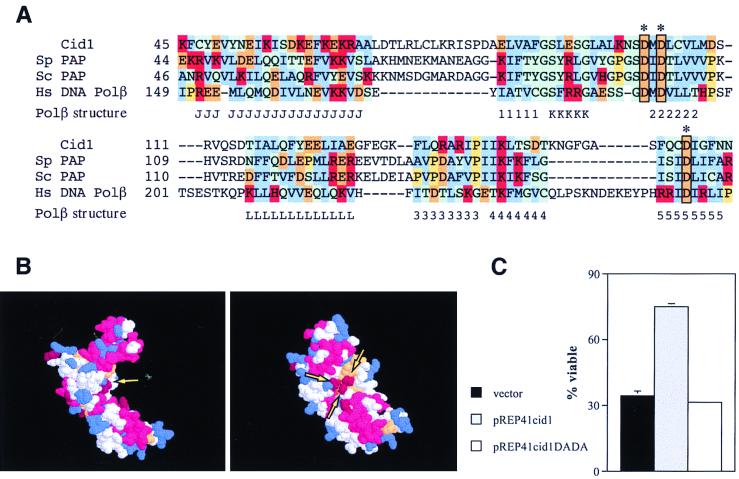
The S-M checkpoint is an intracellular signaling pathway that ensures that mitosis is not initiated in cells undergoing DNA replication. We identified cid1, a novel fission yeast gene, through its ability when overexpressed to confer specific resistance to a combination of hydroxyurea, which inhibits DNA replication, and caffeine, which overrides the S-M checkpoint. Cid1 overexpression also partially suppressed the hydroxyurea sensitivity characteristic of DNA polymerase delta mutants and mutants defective in the "checkpoint Rad" pathway. Cid1 is a member of a family of putative nucleotidyltransferases including budding yeast Trf4 and Trf5, and mutation of amino acid residues predicted to be essential for this activity resulted in loss of Cid1 function in vivo. Two additional Cid1-like proteins play similar but nonredundant checkpoint-signaling roles in fission yeast. Cells lacking Cid1 were found to be viable but specifically sensitive to the combination of hydroxyurea and caffeine and to be S-M checkpoint defective in the absence of Cds1. Genetic data suggest that Cid1 acts in association with Crb2/Rhp9 and through the checkpoint-signaling kinase Chk1 to inhibit unscheduled mitosis specifically when DNA polymerase delta or epsilon is inhibited.
Figures
References
-
- Bahler J, Wu J Q, Longtine M S, Shah N G, McKenzie III A, Steever A B, Wach A, Philippsen P, Pringle J R. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous