

The effects of elevated CO(2) concentrations on cell division rates, growth patterns, and blade anatomy in young wheat plants are modulated by factors related to leaf position, vernalization, and genotype
- PMID: 10759536
- PMCID: PMC58975
- DOI: 10.1104/pp.122.4.1399
The effects of elevated CO(2) concentrations on cell division rates, growth patterns, and blade anatomy in young wheat plants are modulated by factors related to leaf position, vernalization, and genotype
Abstract
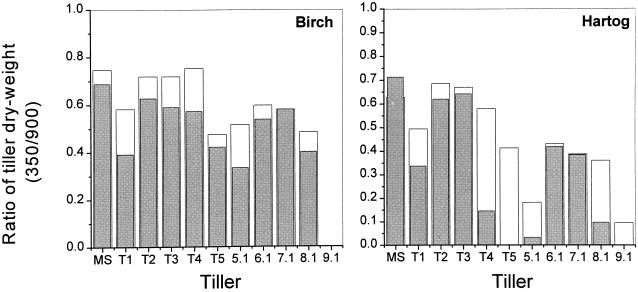
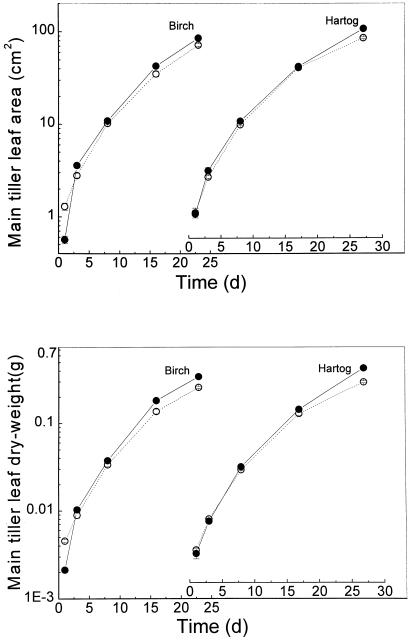
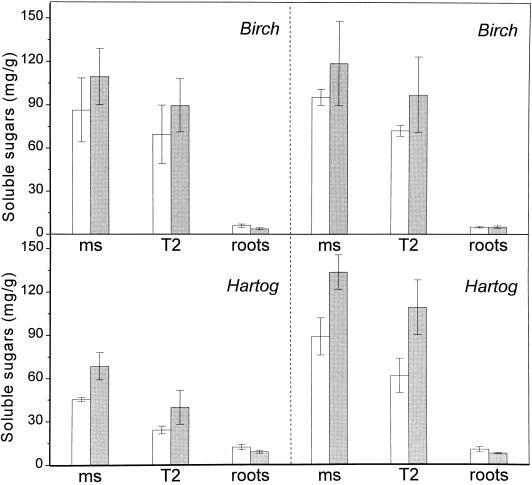
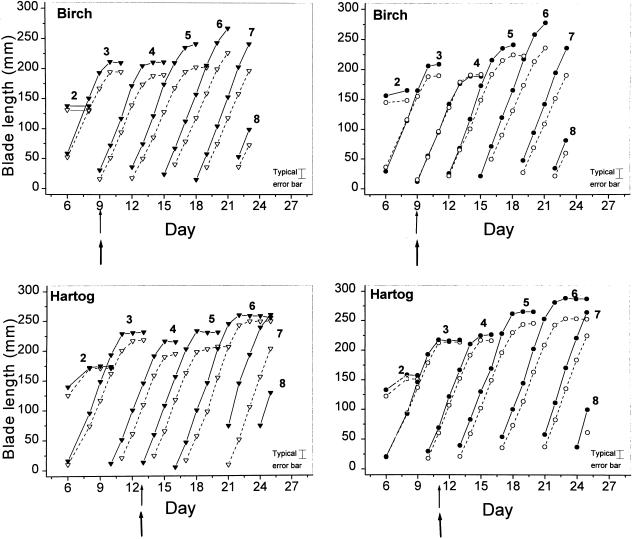
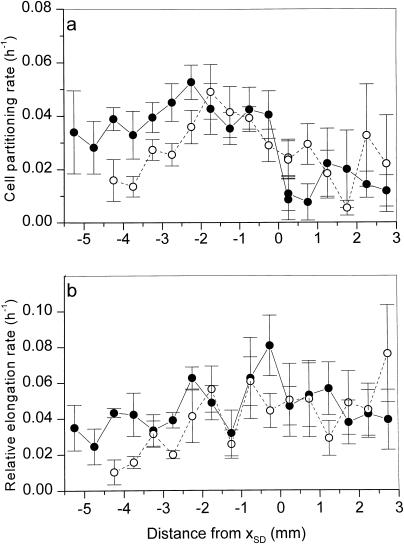
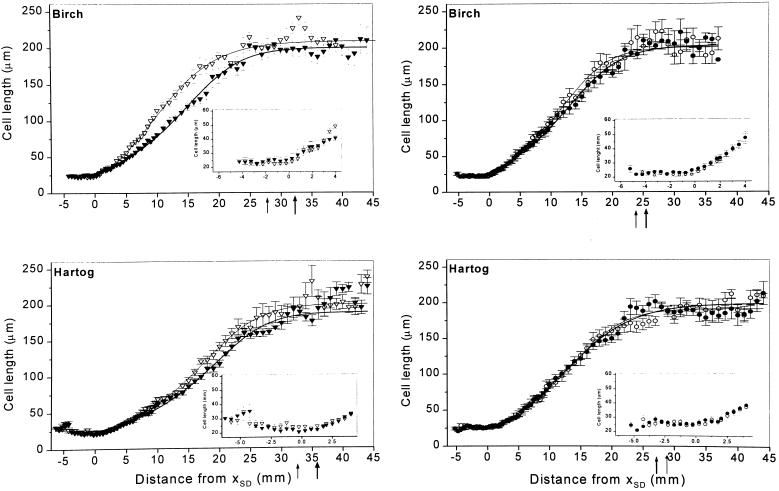
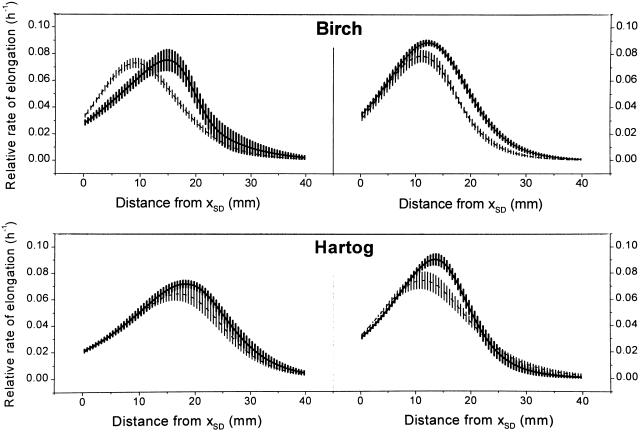
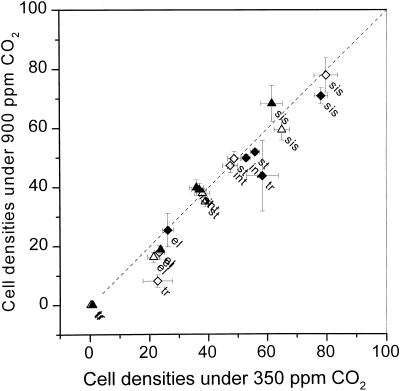
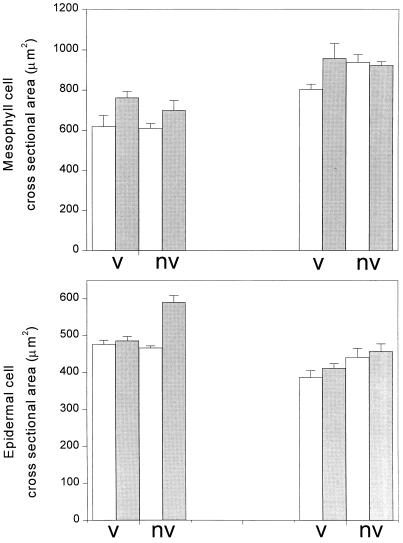
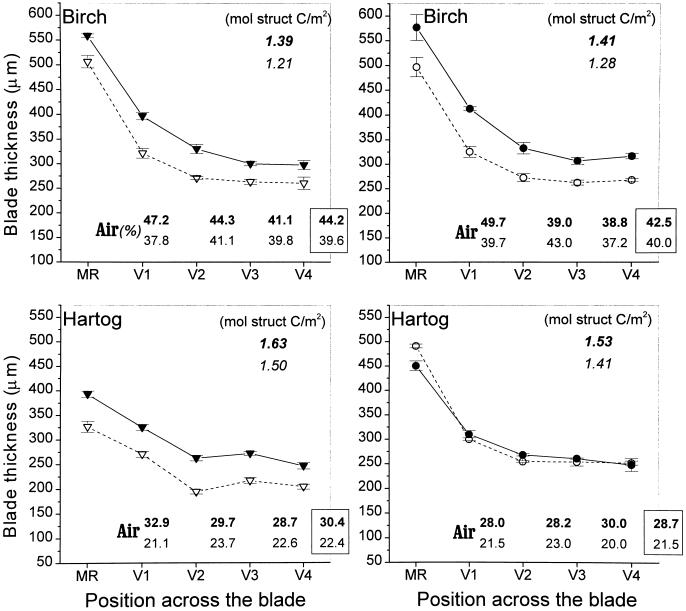
This study demonstrates that elevated [CO(2)] has profound effects on cell division and expansion in developing wheat (Triticum aestivum L.) leaves and on the quantitative integration of these processes in whole-leaf growth kinetics, anatomy, and carbon content. The expression of these effects, however, is modified by intrinsic factors related to genetic makeup and leaf position, and also by exposure to low vernalizing temperatures at germination. Beyond these interactions, leaf developmental responses to elevated [CO(2)] in wheat share several remarkable features that were conserved across all leaves examined. Most significantly: (a) the contribution of [CO(2)] effects on meristem size and activity in driving differences in whole-blade growth kinetics and final dimensions; (b) an anisotropy in cellular growth responses to elevated [CO(2)], with final cell length and expansion in the paradermal plane being highly conserved, even when the rates and duration of cell elongation were modified, while cell cross-sectional areas were increased; (c) tissue-specific effects of elevated [CO(2)], with significant modifications of mesophyll anatomy, including an increased extension of intercellular air spaces and the formation of, on average, one extra cell layer, while epidermal anatomy was mostly unaltered. Our results indicate complex developmental regulations of sugar effects in expanding leaves that are subjected to genetic variation and influenced by environmental cues important in the promotion of floral initiation. They also provide insights into apparently contradictory and inconsistent conclusions of published CO(2) enrichment studies in wheat.
Figures
References
-
- Ballard LAT, Wildman SG. Induction of mitosis by sucrose in excised and attached dormant buds of sunflower (Helianthus annuus L.) Aust J Biol Sci. 1964;17:36–43.
-
- Beemster GTS, Masle J, Williamson RE, Farquhar GD. Effects of soil resistance to root penetration on leaf expansion in wheat (Triticum aestivum L.): kinematic analysis of leaf elongation. J Exp Bot. 1996;47:1663–1678.
-
- Christ RA, Körner C. Responses of shoot and root gas exchange, leaf blade expansion and biomass production to pulses of elevated CO2 in hydroponic wheat. J Exp Bot. 1995;46:1661–1667.
-
- Downton WJ, Björkman O, Pike CS. Consequences of increased atmospheric concentrations of carbon dioxide for growth and photosynthesis of higher plants. In: Pearman GI, editor. Carbon Dioxide and Climate: Australian Research. Australian Academy of Science, Canberra. 1980. pp. 143–151.
-
- Erickson RO, Sax KB. Rates of cell division and cell elongation in the growth of the primary root of Zea mays. Proc Am Philos Soc. 1956;100:499–514.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
