

Auxin regulates the initiation and radial position of plant lateral organs
- PMID: 10760240
- PMCID: PMC139849
- DOI: 10.1105/tpc.12.4.507
Auxin regulates the initiation and radial position of plant lateral organs
Abstract
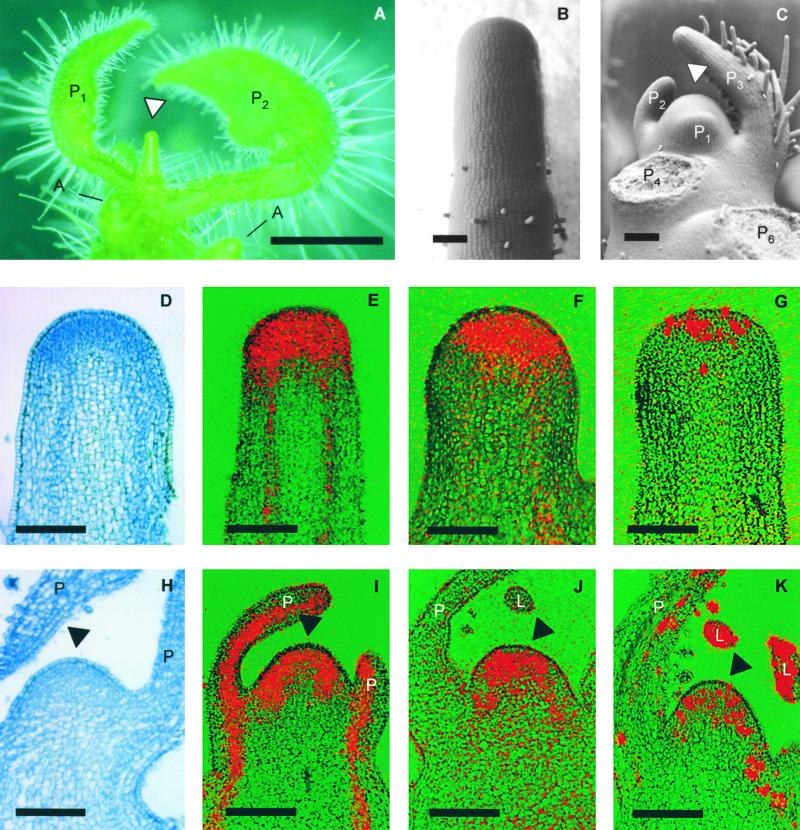
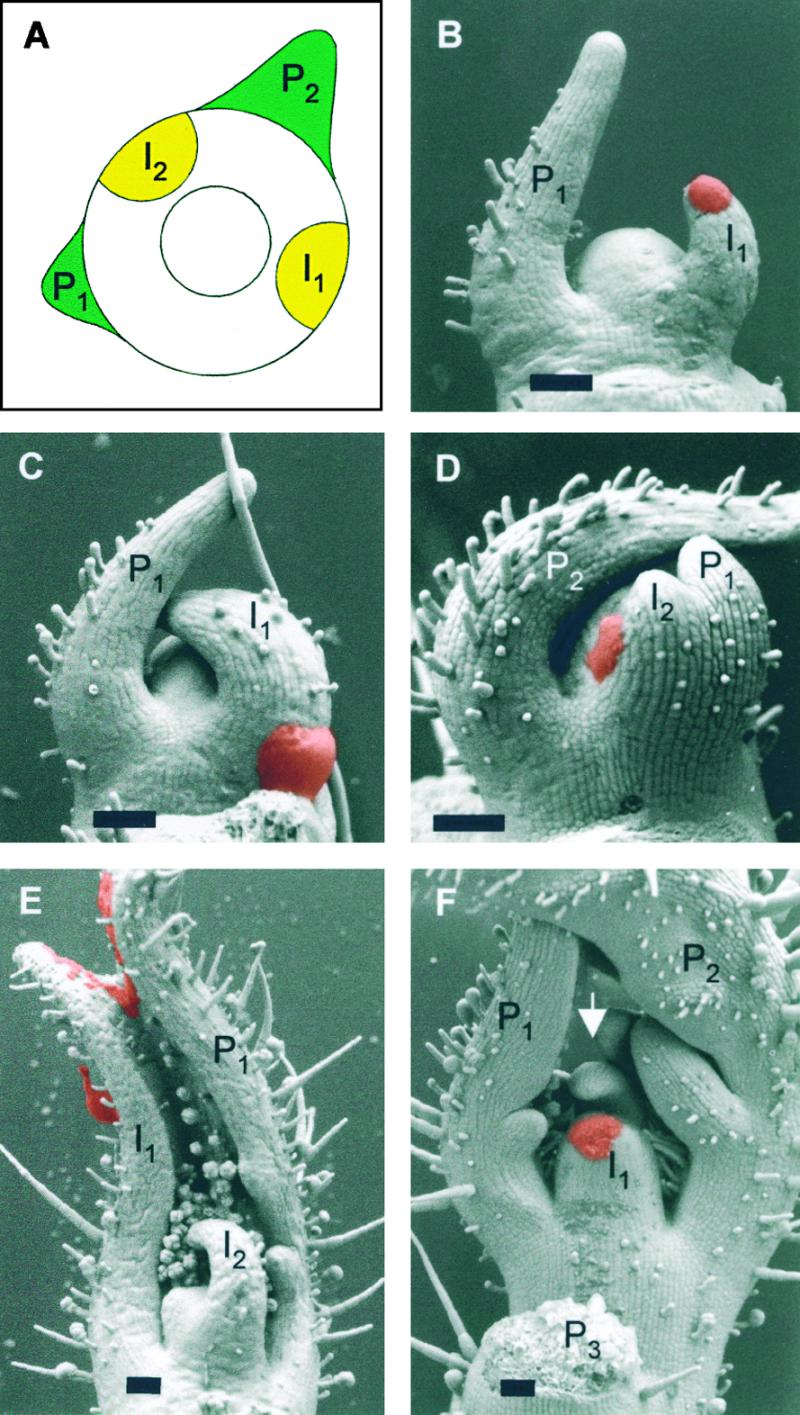
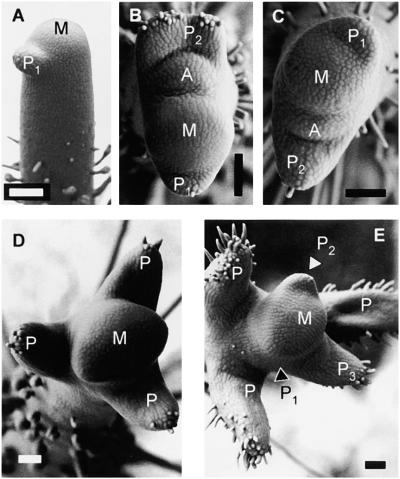
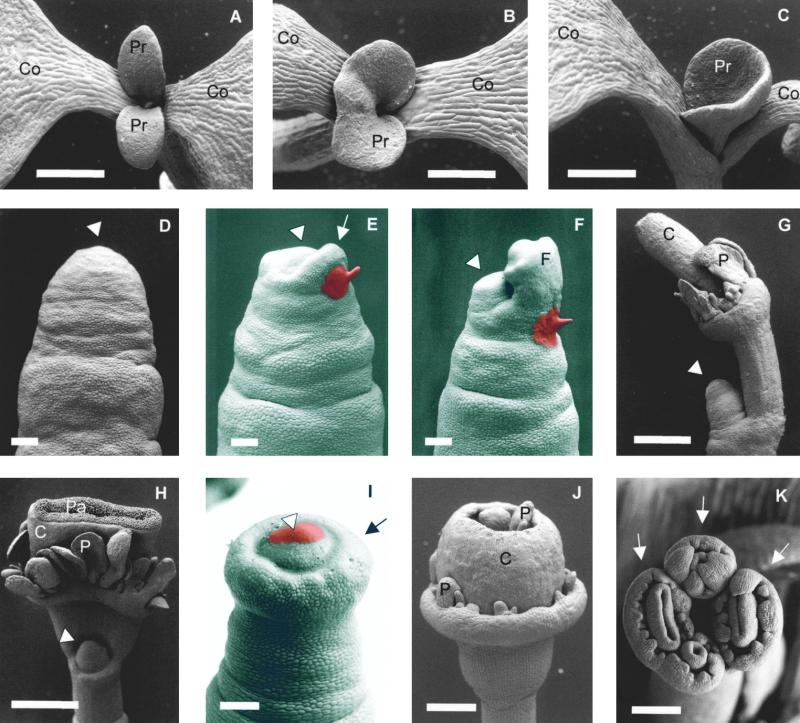
Leaves originate from the shoot apical meristem, a small mound of undifferentiated tissue at the tip of the stem. Leaf formation begins with the selection of a group of founder cells in the so-called peripheral zone at the flank of the meristem, followed by the initiation of local growth and finally morphogenesis of the resulting bulge into a differentiated leaf. Whereas the mechanisms controlling the switch between meristem propagation and leaf initiation are being identified by genetic and molecular analyses, the radial positioning of leaves, known as phyllotaxis, remains poorly understood. Hormones, especially auxin and gibberellin, are known to influence phyllotaxis, but their specific role in the determination of organ position is not clear. We show that inhibition of polar auxin transport blocks leaf formation at the vegetative tomato meristem, resulting in pinlike naked stems with an intact meristem at the tip. Microapplication of the natural auxin indole-3-acetic acid (IAA) to the apex of such pins restores leaf formation. Similarly, exogenous IAA induces flower formation on Arabidopsis pin-formed1-1 inflorescence apices, which are blocked in flower formation because of a mutation in a putative auxin transport protein. Our results show that auxin is required for and sufficient to induce organogenesis both in the vegetative tomato meristem and in the Arabidopsis inflorescence meristem. In this study, organogenesis always strictly coincided with the site of IAA application in the radial dimension, whereas in the apical-basal dimension, organ formation always occurred at a fixed distance from the summit of the meristem. We propose that auxin determines the radial position and the size of lateral organs but not the apical-basal position or the identity of the induced structures.
Figures

References
-
- Bedesem, P.P. (1958). Histogenetic effects of 2,3,5-triiodobenzoic acid on the shoot apices and leaf primordia of tomato. Bull. Torrey Bot. Club 85, 434–472.
-
- Brandstätter, J., Rossbach, C., and Theres, K. (1994). The pattern of histone H4 expression in the tomato shoot apex changes during development. Planta 192, 69–74. - PubMed
-
- Callos, J.D., and Medford, J.I. (1994). Organ positions and pattern formation in the shoot apex. Plant J. 6, 1–7.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
