

DNA repair protein Ku80 suppresses chromosomal aberrations and malignant transformation
- PMID: 10761921
- PMCID: PMC4721590
- DOI: 10.1038/35006670
DNA repair protein Ku80 suppresses chromosomal aberrations and malignant transformation
Abstract
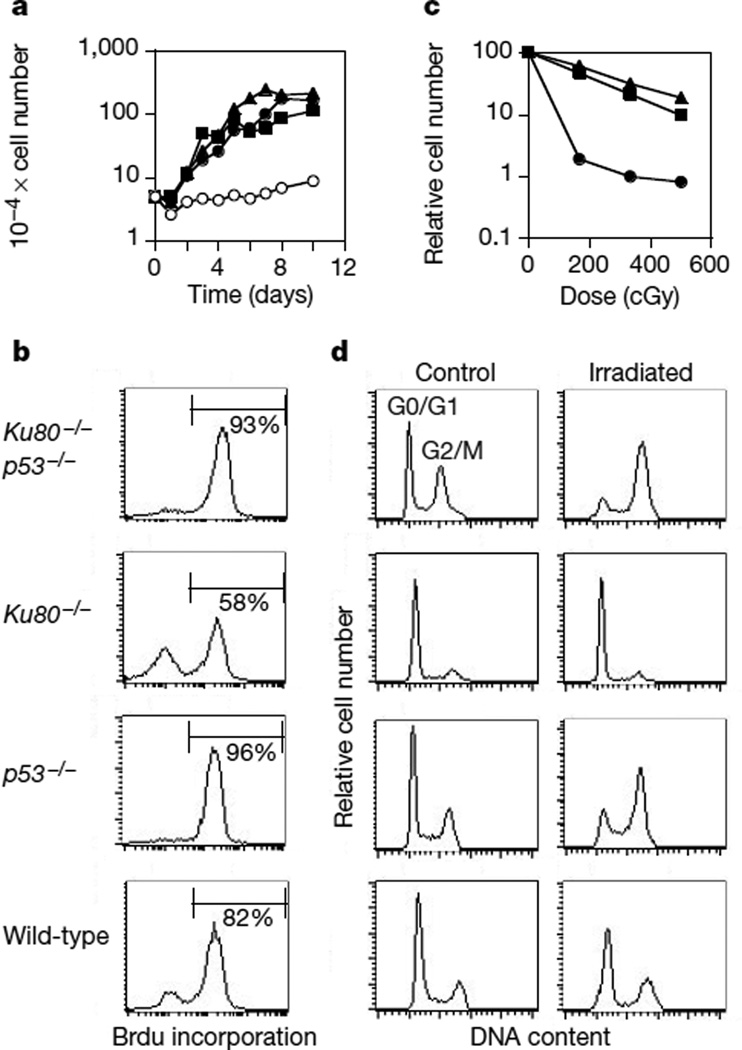
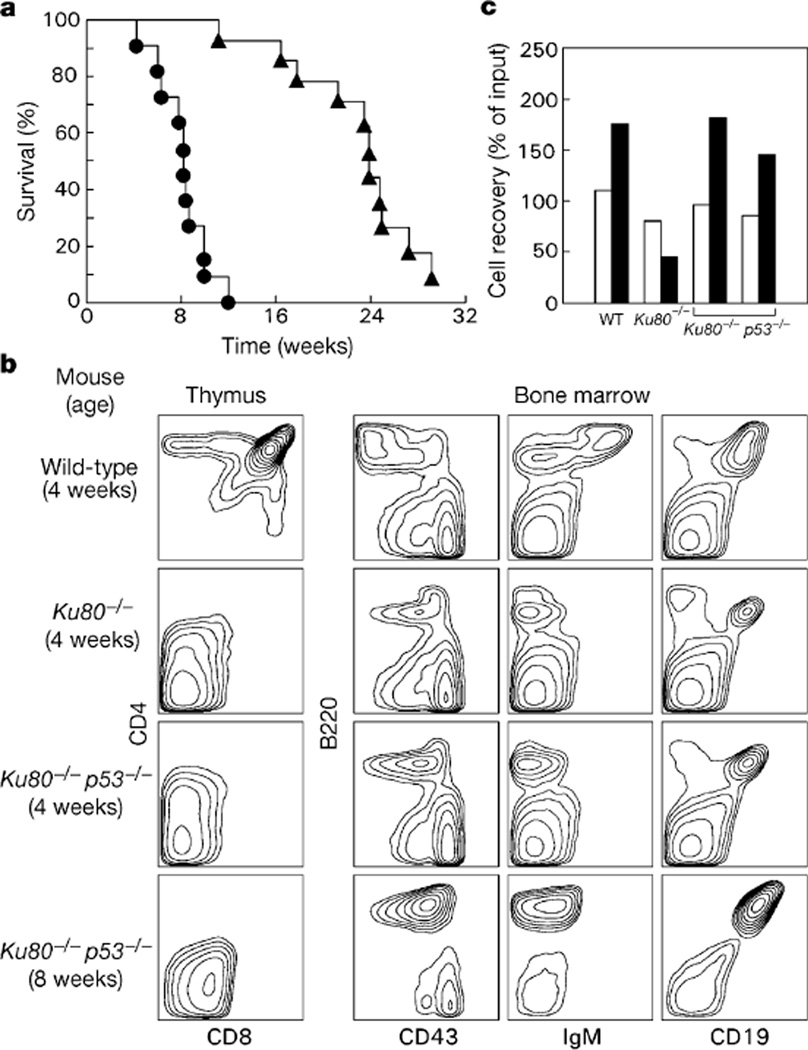
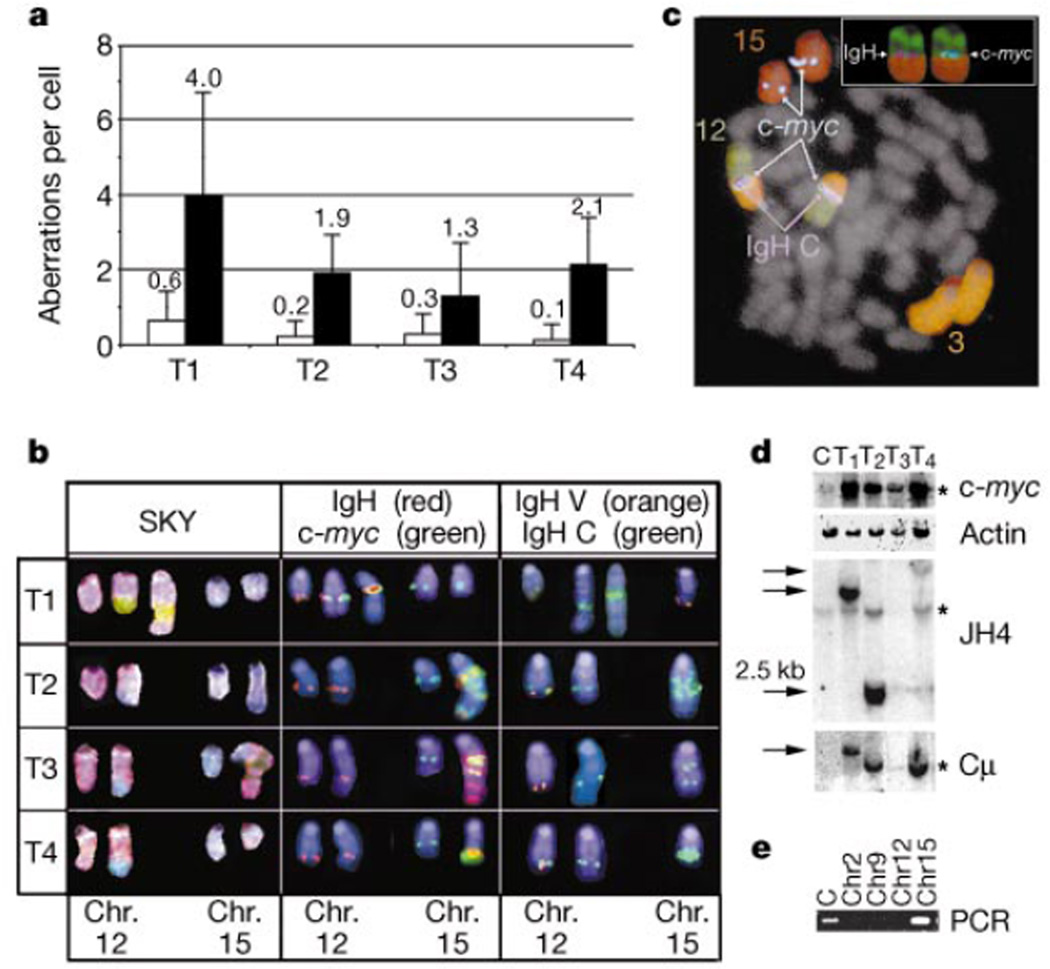
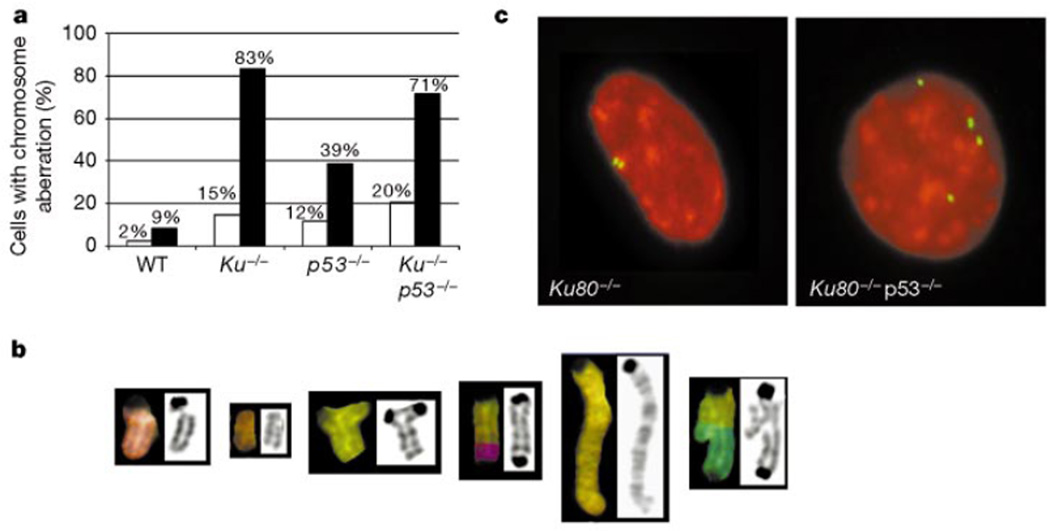
Cancer susceptibility genes have been classified into two groups: gatekeepers and caretakers. Gatekeepers are genes that control cell proliferation and death, whereas caretakers are DNA repair genes whose inactivation leads to genetic instability. Abrogation of both caretaker and gatekeeper function markedly increases cancer susceptibility. Although the importance of Ku80 in DNA double-strand break repair is well established, neither Ku80 nor other components of the non-homologous end-joining pathway are known to have a caretaker role in maintaining genomic stability. Here we show that mouse cells deficient for Ku80 display a marked increase in chromosomal aberrations, including breakage, translocations and aneuploidy. Despite the observed chromosome instabilities, Ku80-/- mice have only a slightly earlier onset of cancer. Loss of p53 synergizes with Ku80 to promote tumorigenesis such that all Ku80-/- p53-/- mice succumb to disseminated pro-B-cell lymphoma before three months of age. Tumours result from a specific set of chromosomal translocations and gene amplifications involving IgH and c-Myc, reminiscent of Burkitt's lymphoma. We conclude that Ku80 is a caretaker gene that maintains the integrity of the genome by a mechanism involving the suppression of chromosomal rearrangements.
Figures
Comment in
-
New guardians of the genome.Nature. 2000 Apr 20;404(6780):823-5. doi: 10.1038/35009180. Nature. 2000. PMID: 10786775 No abstract available.
References
-
- Kinzler KW, Vogelstein B. Gatekeepers and caretakers. Nature. 1997;386:761–763. - PubMed
-
- Nussenzweig A, et al. Requirement for Ku80 in growth and immunoglobulin V(D)J recombination. Nature. 1996;382:551–555. - PubMed
-
- Zhu C, Bogue MA, Lim DS, Hasty P, Roth DB. Ku86-deficient mice exhibit severe combined immunodeficiency and defective processing of V(D)J recombination intermediates. Cell. 1996;86:379–389. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
