

Retinal innervation of calbindin-D28K cells in the hamster suprachiasmatic nucleus: ultrastructural characterization
- PMID: 10762028
- PMCID: PMC3281760
- DOI: 10.1177/074873040001500204
Retinal innervation of calbindin-D28K cells in the hamster suprachiasmatic nucleus: ultrastructural characterization
Abstract
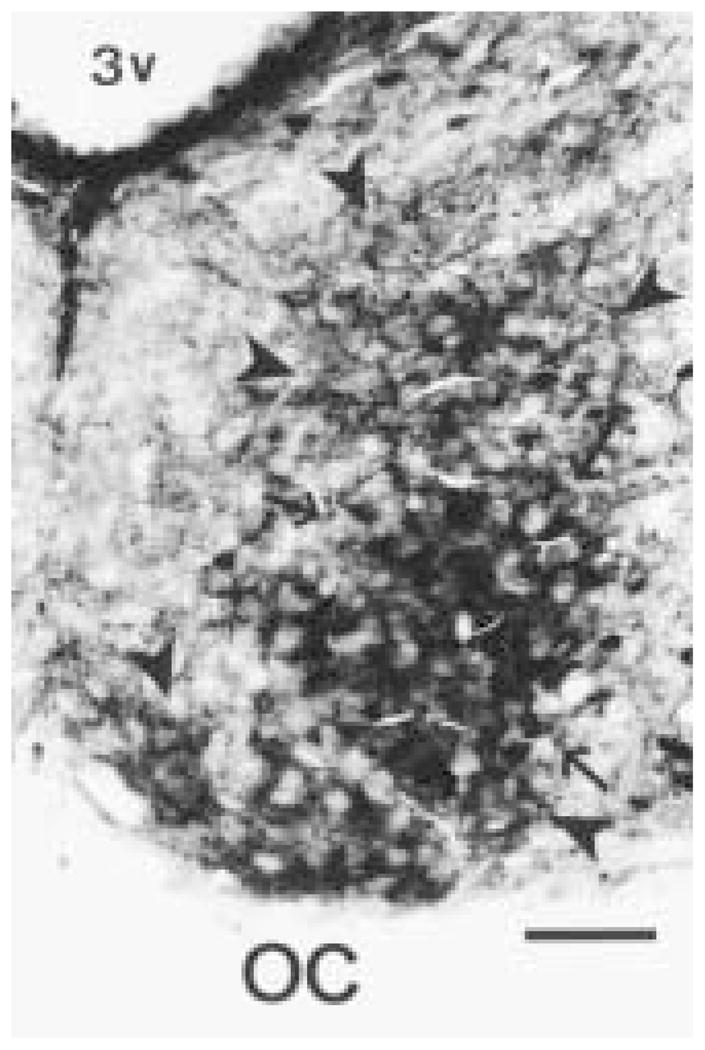
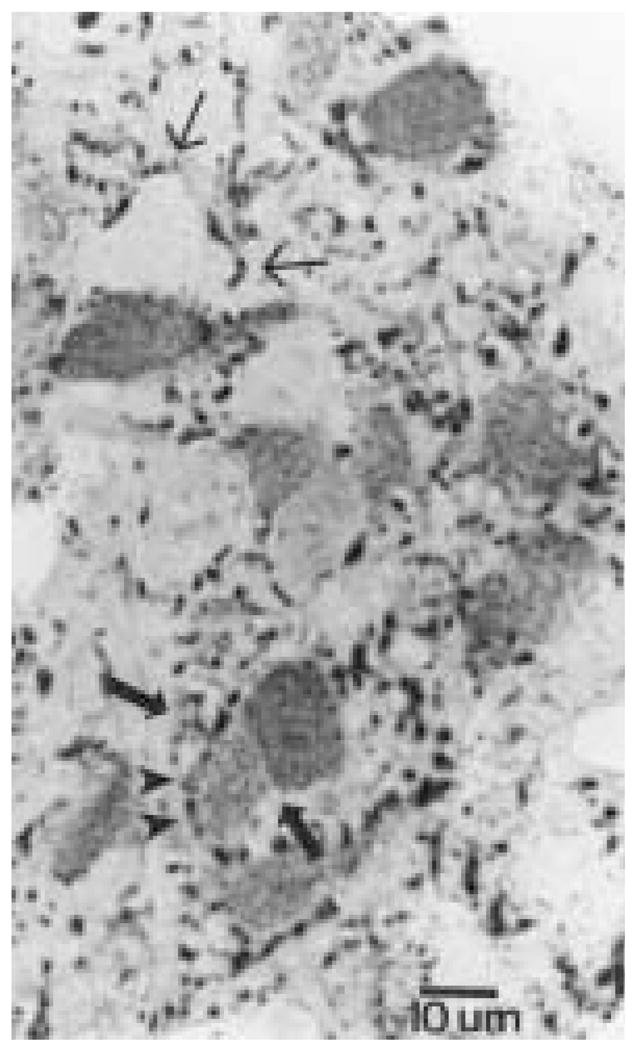
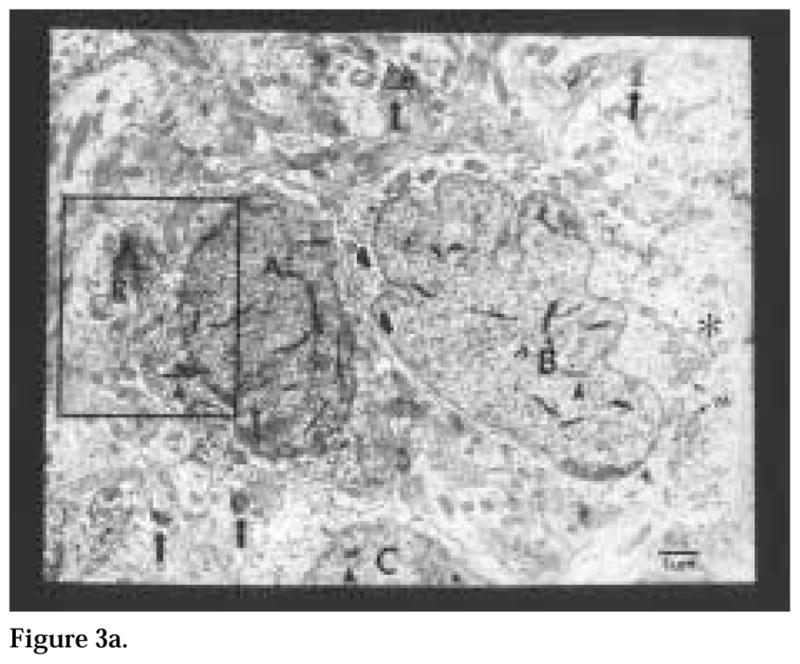
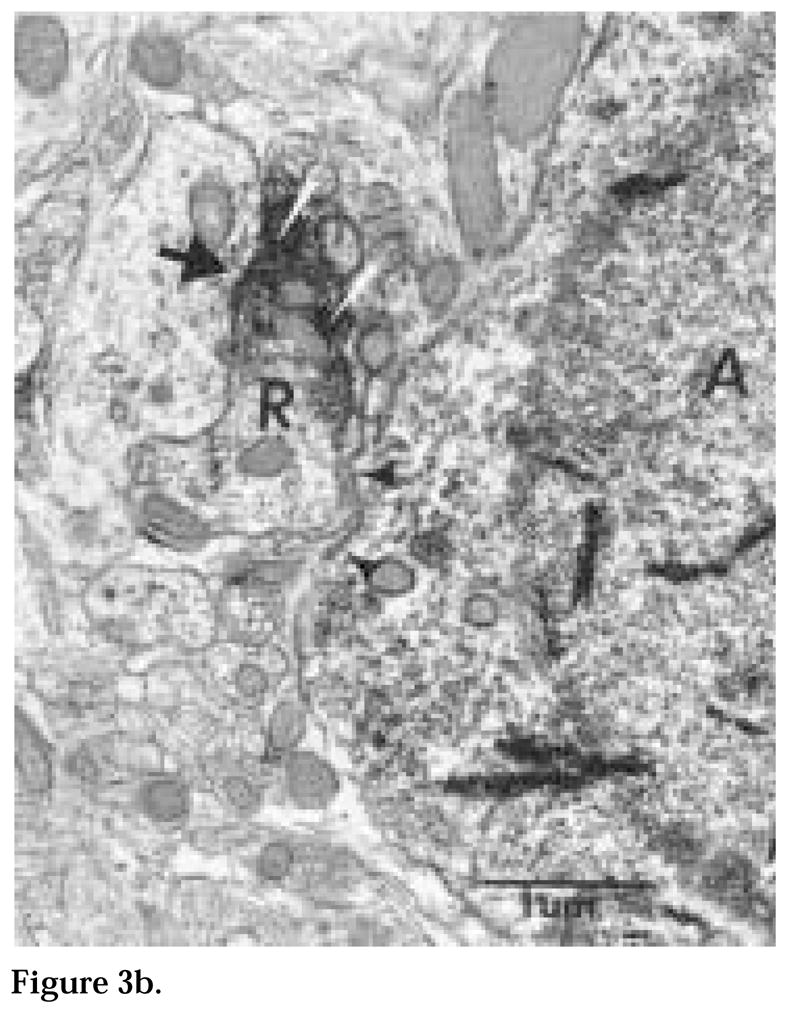
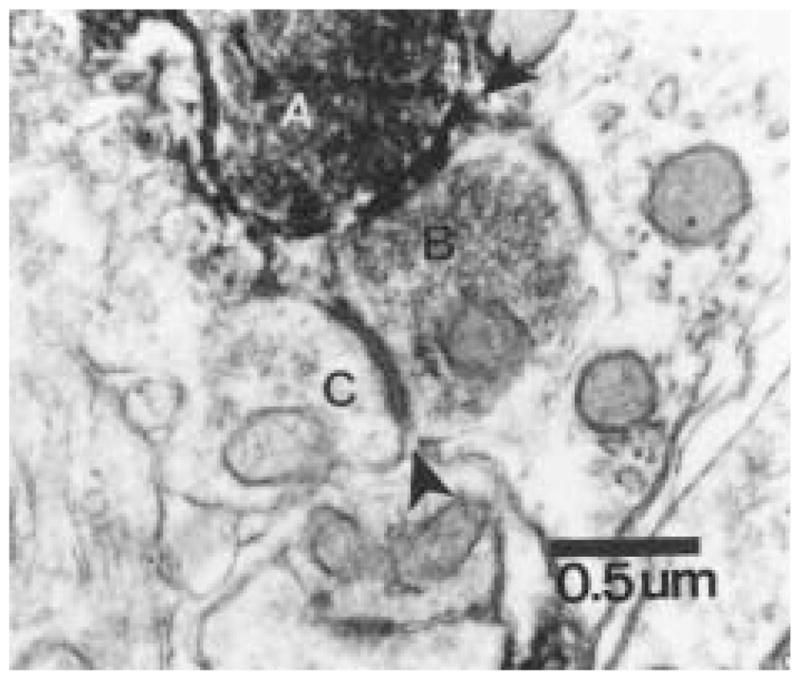
The authors have described a subregion of the hamster hypothalamic suprachiasmatic nucleus (SCN) containing cells that are immunopositive for the cytosolic calcium-binding protein, Calbindin-D28K (CaBP). Several lines of evidence indicate that this region may constitute the site of the pacemaker cells that are responsible for the regulation of circadian locomotor rhythms. First, 79% of the CaBP-immunoreactive (ir) neurons express Fos in response to photic stimulation, indicating that they are close to or part of the input pathway to pacemakers. Second, at the light microscopy level, retinal terminals innervate the CaBP subnucleus. Finally, destruction of this subnucleus renders animals arrhythmic in locomotor activity. In this study, the authors examined the ultrastructural relationship between cholera toxin (CTbeta) labeled retinal fibers and the CaBP-ir subregion within the hamster SCN. CTbeta-ir retinal terminals make primarily axo-somatic, symmetric, synaptic contacts with CaBP-ir perikarya. In addition, retinal terminals form synapses with CaBP processes as well as with unidentified profiles. There are also complex interactions between retinal terminals, CaBP perikarya, and unidentified profiles. Given that axo-somatic synaptic input has a more potent influence on a cell's electrical activity than does axo-dendritic synaptic input, cells of the CaBP subregion of the SCN are ideally suited to respond rapidly to photic stimulation to reset circadian pacemakers.
Figures
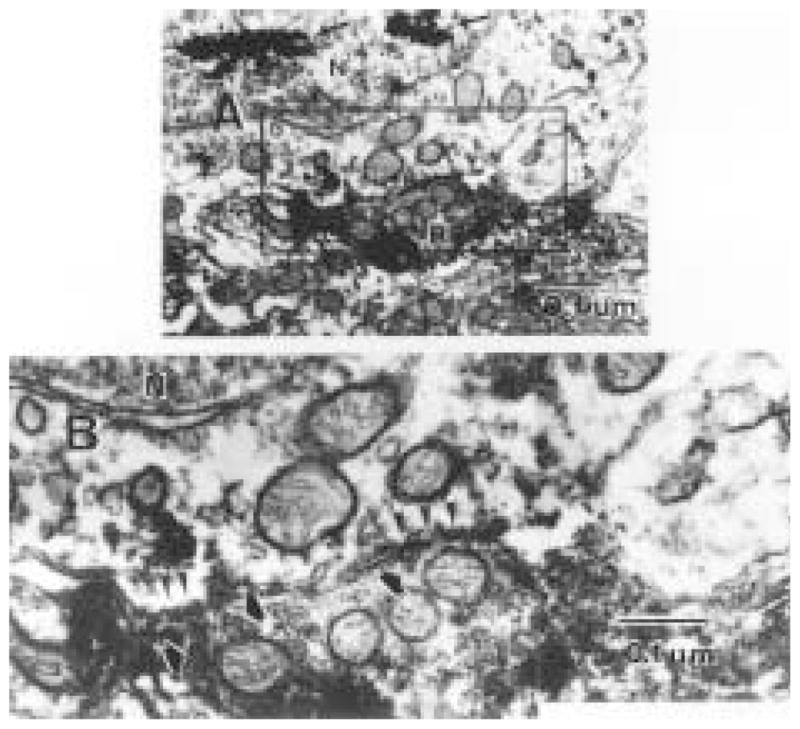
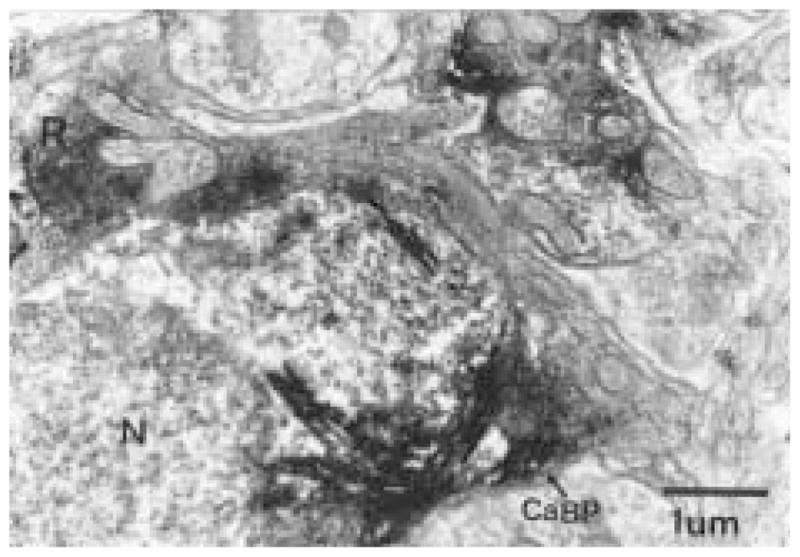
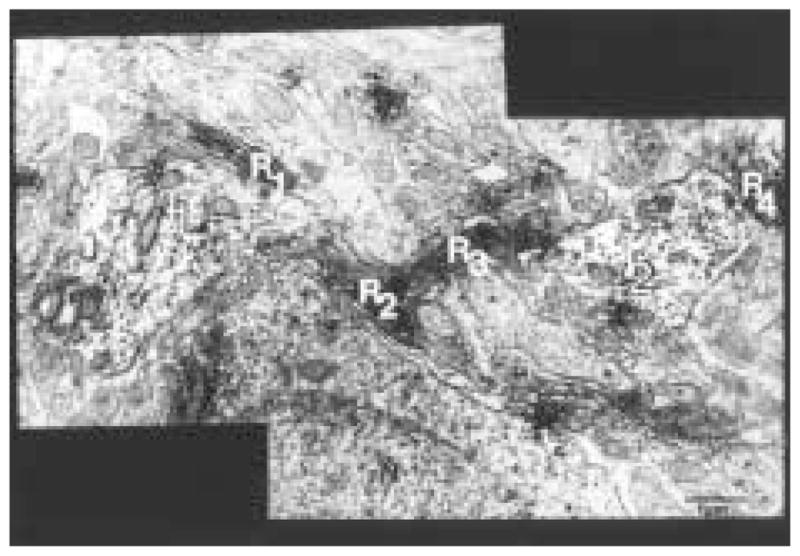
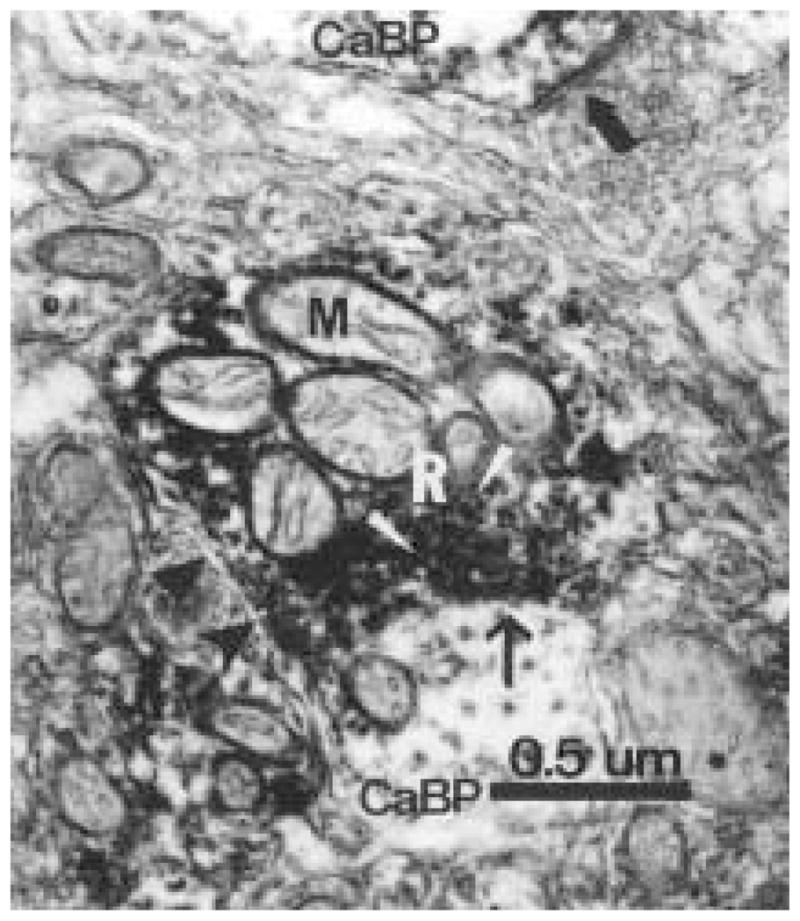
References
-
- Abrahamson EE, Moore RY. The organization of the mouse suprachiasmatic nucleus (SCN): Immunocytochemical analysis of retinal innervation and cell and fiber distribution. Soc Res Biol Rhythms Abstr. 1998;6:83.
-
- Aïoun J, Chambille I, Peytevin J, Martinet L. Neurons containing gastrin releasing peptide and vasoactive intestinal polypeptide are involved in the reception of the photic signal in the suprachiasmatic nucleus of the Syrian hamster: An immunocytochemical ultrastructural study. Cell Tissue Res. 1998;291:231–253. - PubMed
-
- Baimbridge KG, Celio MR, Rogers JH. Calcium-binding proteins in the nervous system. Trends Neurosci. 1992;15:303–308. - PubMed
-
- Bryant DN, LeSauter J, Silver R, Romero M-T. Retinal synapses on calbindin-ir cells in the hamster suprachiasmatic nucleus: A double label immunoelectron microscopy study. Soc Neurosci Abstr. 1996;22:1140.
-
- Burke RE. Spinal cord: Ventral horn. In: Shepherd GM, editor. The Synaptic Organization of the Brain. Oxford University Press; New York: 1998. pp. 77–120.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
