

Action of RNase II and polynucleotide phosphorylase against RNAs containing stem-loops of defined structure
- PMID: 10762241
- PMCID: PMC111303
- DOI: 10.1128/JB.182.9.2422-2427.2000
Action of RNase II and polynucleotide phosphorylase against RNAs containing stem-loops of defined structure
Abstract
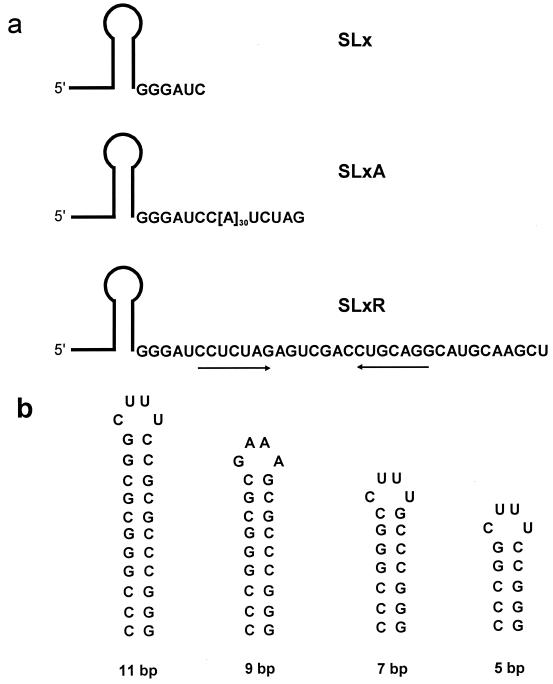
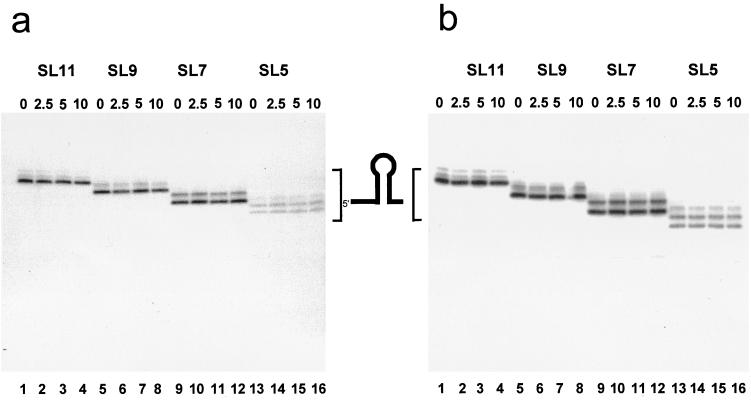
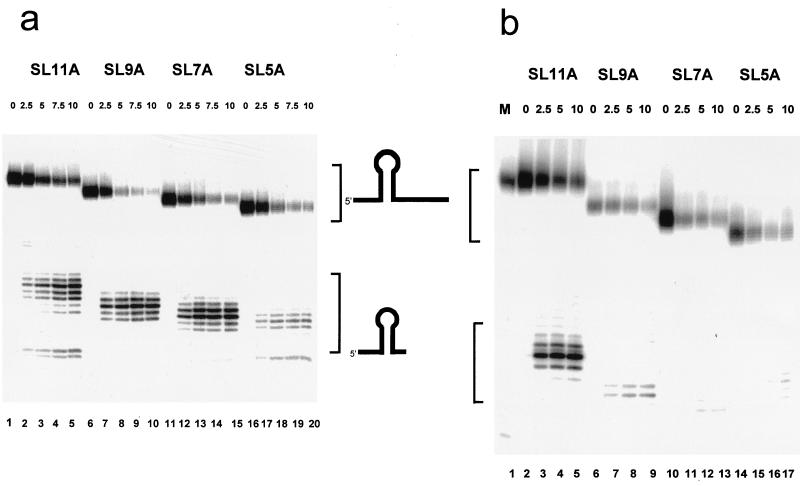
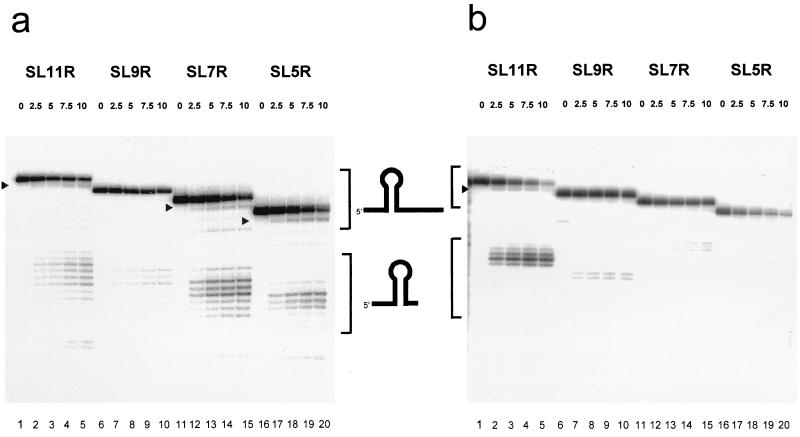
The 3'-->5' exoribonucleases, RNase II and polynucleotide phosphorylase (PNPase), play an essential role in degrading fragments of mRNA generated by prior cleavages by endonucleases. We have assessed the ability of small RNA substrates containing defined stem-loop structures and variable 3' extensions to impede the exonucleolytic activity of these enzymes. We find that stem-loops containing five G-C base pairs do not block either enzyme; in contrast, more stable stem-loops of 7, 9, or 11 bp block the processive action of both enzymes. Under conditions where enzyme activity is limiting, both enzymes stall and dissociate from their substrates six to nine residues, on average, from the base of a stable stem-loop structure. Our data provide a clear mechanistic explanation for the previous observation that RNase II and PNPase behave as functionally redundant.
Figures
Similar articles
-
Different specificities of ribonuclease II and polynucleotide phosphorylase in 3'mRNA decay.Biochimie. 1990 Nov;72(11):771-7. doi: 10.1016/0300-9084(90)90186-k. Biochimie. 1990. PMID: 2085542 Review.
-
Different specificities of ribonuclease II and polynucleotide phosphorylase in 3'mRNA decay.Biochimie. 1991 May;73(5):543-9. doi: 10.1016/0300-9084(91)90021-r. Biochimie. 1991. PMID: 1764498 Review.
-
The Phosphorolytic Exoribonucleases Polynucleotide Phosphorylase and RNase PH Stabilize sRNAs and Facilitate Regulation of Their mRNA Targets.J Bacteriol. 2016 Nov 18;198(24):3309-3317. doi: 10.1128/JB.00624-16. Print 2016 Dec 15. J Bacteriol. 2016. PMID: 27698082 Free PMC article.
-
mRNA degradation by processive 3'-5' exoribonucleases in vitro and the implications for prokaryotic mRNA decay in vivo.J Mol Biol. 1991 Sep 5;221(1):81-95. J Mol Biol. 1991. PMID: 1920421
-
Decay of the IS10 antisense RNA by 3' exoribonucleases: evidence that RNase II stabilizes RNA-OUT against PNPase attack.Mol Microbiol. 1994 Sep;13(6):1133-42. doi: 10.1111/j.1365-2958.1994.tb00504.x. Mol Microbiol. 1994. PMID: 7531807
Cited by
-
Polynucleotide phosphorylase is a global regulator of virulence and persistency in Salmonella enterica.Proc Natl Acad Sci U S A. 2002 Jun 25;99(13):8784-9. doi: 10.1073/pnas.132047099. Epub 2002 Jun 18. Proc Natl Acad Sci U S A. 2002. PMID: 12072563 Free PMC article.
-
The roles of individual domains of RNase R in substrate binding and exoribonuclease activity. The nuclease domain is sufficient for digestion of structured RNA.J Biol Chem. 2009 Jan 2;284(1):486-494. doi: 10.1074/jbc.M806468200. Epub 2008 Nov 11. J Biol Chem. 2009. PMID: 19004832 Free PMC article.
-
Polynucleotide phosphorylase: Not merely an RNase but a pivotal post-transcriptional regulator.PLoS Genet. 2018 Oct 11;14(10):e1007654. doi: 10.1371/journal.pgen.1007654. eCollection 2018 Oct. PLoS Genet. 2018. PMID: 30307990 Free PMC article. Review.
-
Nonspherical Coacervate Shapes in an Enzyme-Driven Active System.Langmuir. 2020 Mar 3;36(8):1956-1964. doi: 10.1021/acs.langmuir.9b02719. Epub 2020 Feb 17. Langmuir. 2020. PMID: 31995710 Free PMC article.
-
A newly identified duplex RNA unwinding activity of archaeal RNase J depends on processive exoribonucleolysis coupled steric occlusion by its structural archaeal loops.RNA Biol. 2020 Oct;17(10):1480-1491. doi: 10.1080/15476286.2020.1777379. Epub 2020 Jun 18. RNA Biol. 2020. PMID: 32552320 Free PMC article.
References
-
- Belasco J G. mRNA degradation in prokaryotic cells: an overview. In: Belasco J G, Brawerman G, editors. Control of messenger RNA stability. San Diego, Calif: Academic Press; 1993. pp. 3–12.
-
- Blum E, Carpousis A J, Higgins C F. Polyadenylation promotes degradation of 3′-structured RNA by the Escherichia coli mRNA degradosome in vitro. J Biol Chem. 1999;274:4009–4016. - PubMed
-
- Braun F, Hajnsdorf E, Régnier P. Polynucleotide phosphorylase is required for the rapid degradation of the RNase E-processed rpsO mRNA of Escherichia coli devoid of its 3′ hairpin. Mol Microbiol. 1996;19:997–1005. - PubMed
-
- Cannistraro V J, Kennell D. The processive reaction mechanism of ribonuclease II. J Mol Biol. 1994;243:930–943. - PubMed
-
- Carpousis A J, Van Houwe G, Ehretsmann C, Krisch H M. Copurification of E. coli RNase E and PNPase: evidence for a specific association between two enzymes important in RNA processing and degradation. Cell. 1994;76:889–900. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
