

Effects of bfp mutations on biogenesis of functional enteropathogenic Escherichia coli type IV pili
- PMID: 10762251
- PMCID: PMC111313
- DOI: 10.1128/JB.182.9.2498-2506.2000
Effects of bfp mutations on biogenesis of functional enteropathogenic Escherichia coli type IV pili
Abstract
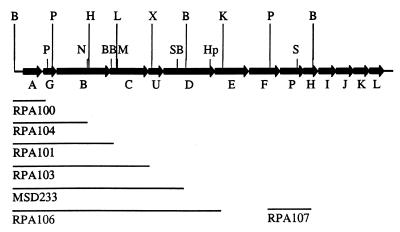
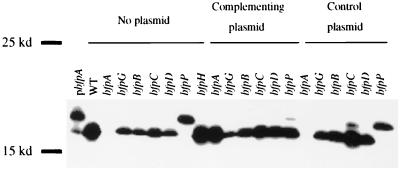
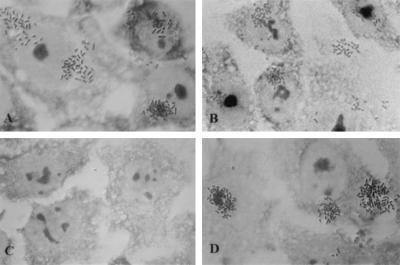
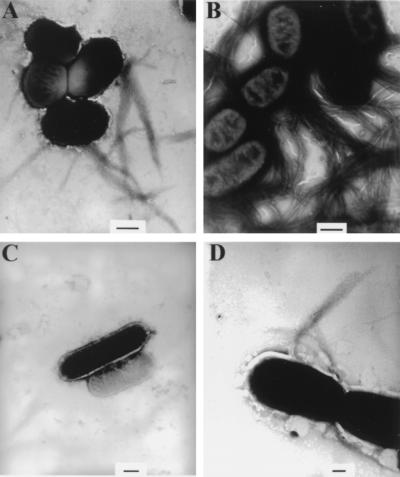
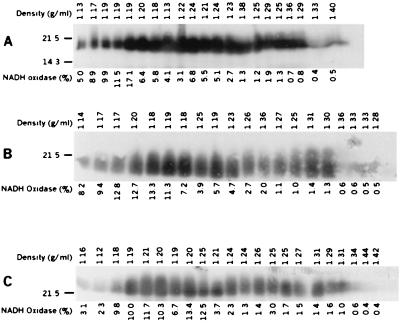
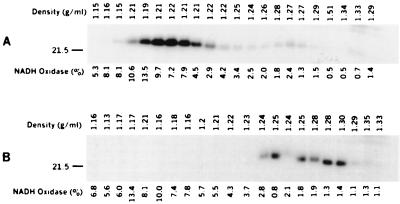
Enteropathogenic Escherichia coli expresses a type IV fimbria known as the bundle-forming pilus (BFP) that is required for autoaggregation and localized adherence (LA) to host cells. A cluster of 14 genes is sufficient to reconstitute BFP biogenesis in a laboratory strain of E. coli. We have undertaken a systematic mutagenesis of the individual genes to determine the effect of each mutation on BFP biogenesis and LA. Here we report the construction and analysis of nonpolar mutations in six genes of the bfp cluster, bfpG, bfpB, bfpC, bfpD, bfpP, and bfpH, as well as the further analysis of a previously described bfpA mutant strain that is unable to express bundlin, the pilin protein. We found that mutations in bfpB, which encodes an outer membrane protein; bfpD, which encodes a putative nucleotide-binding protein; and bfpG and bfpC, which do not have sequence homologues in other type IV pilus systems, do not affect prebundlin expression or processing but block both BFP biogenesis and LA. The mutation in bfpP, the prepilin peptidase gene, does not affect prebundlin expression but blocks signal sequence cleavage of prebundlin, BFP biogenesis, and LA. The mutation in bfpH, which is predicted to encode a lytic transglycosylase, has no effect on prebundlin expression, prebundlin processing, BFP biogenesis, or LA. For each mutant for which altered phenotypes were detected, complementation with a plasmid containing the corresponding wild-type allele restored the wild-type phenotypes. We also found that association of prebundlin or bundlin with sucrose density flotation gradient fractions containing both inner and outer membrane proteins does not require any accessory proteins. These studies indicate that many bfp gene products are required for biogenesis of functional type IV pili but that mutations in the individual genes do not lead to the identification of new phases of pilus assembly.
Figures
References
-
- Alm R A, Hallinan J P, Watson A A, Mattick J S. Fimbrial biogenesis genes of Pseudomonas aeruginosa: PilW and pilX increase the similarity of type 4 fimbriae to the GSP protein-secretion systems and pilY1 encodes a gonococcal PilC homologue. Mol Microbiol. 1996;22:161–173. - PubMed
-
- Alm R A, Mattick J S. Identification of a gene, pilV, required for type 4 fimbrial biogenesis in Pseudomonas aeruginosa, whose product possesses a pre-pilin-like leader sequence. Mol Microbiol. 1995;16:485–496. - PubMed
-
- Alm R A, Mattick J S. Genes involved in the biogenesis and function of type-4 fimbriae in Pseudomonas aeruginosa. Gene. 1997;192:89–98. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
