

Kinetics and substrate specificity of membrane-reconstituted peptide transporter DtpT of Lactococcus lactis
- PMID: 10762255
- PMCID: PMC111317
- DOI: 10.1128/JB.182.9.2530-2535.2000
Kinetics and substrate specificity of membrane-reconstituted peptide transporter DtpT of Lactococcus lactis
Abstract
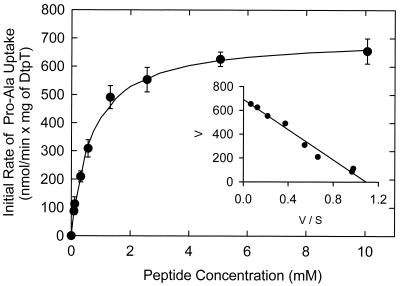
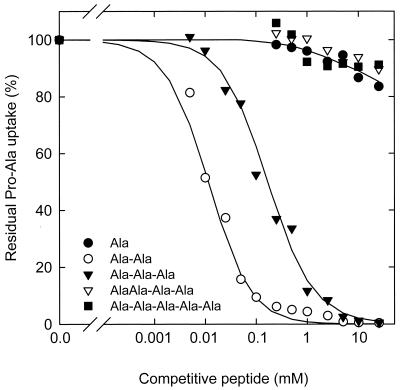
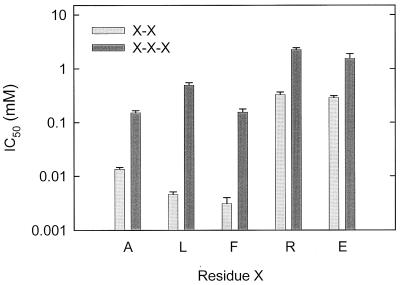
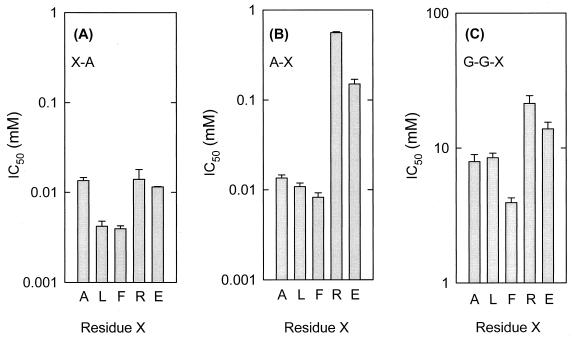
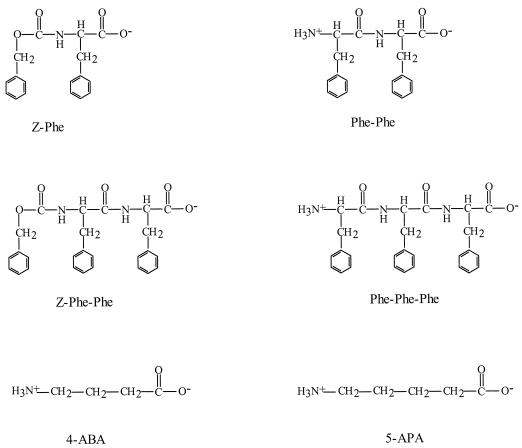
The peptide transport protein DtpT of Lactococcus lactis was purified and reconstituted into detergent-destabilized liposomes. The kinetics and substrate specificity of the transporter in the proteoliposomal system were determined, using Pro-[(14)C]Ala as a reporter peptide in the presence of various peptides or peptide mimetics. The DtpT protein appears to be specific for di- and tripeptides, with the highest affinities for peptides with at least one hydrophobic residue. The effect of the hydrophobicity, size, or charge of the amino acid was different for the amino- and carboxyl-terminal positions of dipeptides. Free amino acids, omega-amino fatty acid compounds, or peptides with more than three amino acid residues do not interact with DtpT. For high-affinity interaction with DtpT, the peptides need to have free amino and carboxyl termini, amino acids in the L configuration, and trans-peptide bonds. Comparison of the specificity of DtpT with that of the eukaryotic homologues PepT(1) and PepT(2) shows that the bacterial transporter is more restrictive in its substrate recognition.
Figures
References
-
- Basrai M A, Lubkowitz M A, Perry J R, Miller D, Krainer E, Naider F, Becker J M. Cloning of a Candida albicans peptide transport gene. Microbiology. 1995;141:1147–1156. - PubMed
-
- Börner V, Fei Y J, Hartrodt B, Ganapathy V, Leibach F H, Neubert K, Brandsch M. Transport of amino acid aryl amides by the intestinal H+/peptide cotransport system, PEPT1. Eur J Biochem. 1998;255:698–702. - PubMed
-
- Chen X Z, Zhu T, Smith D E, Hediger M A. Stoichiometry and kinetics of the high-affinity H+-coupled peptide transporter PepT2. J Biol Chem. 1999;274:2773–2779. - PubMed
-
- Daniel H, Morse E L, Adibi S A. Determinants of substrate affinity for the oligopeptide/H+ symporter in the renal brush border membrane. J Biol Chem. 1992;267:9565–9573. - PubMed
-
- Detmers F J M, Kunji E R S, Lanfermeijer F C, Poolman B, Konings W N. Kinetics and specificity of peptide uptake by the oligopeptide transport system of Lactococcus lactis. Biochemistry. 1998;37:16671–16679. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
