

Tetanus toxin fragment C expressed in live Salmonella vaccines enhances antibody responses to its fusion partner Schistosoma haematobium glutathione S-transferase
- PMID: 10768937
- PMCID: PMC97452
- DOI: 10.1128/IAI.68.5.2503-2512.2000
Tetanus toxin fragment C expressed in live Salmonella vaccines enhances antibody responses to its fusion partner Schistosoma haematobium glutathione S-transferase
Abstract
Tetanus toxoid has been used widely as an adjuvant. The atoxic fragment C from tetanus toxin (TetC) is potently immunogenic when expressed in Salmonella vaccine strains and has been used as a fusion partner for antigens (Ag). However, there has been no formal comparison of the immunomodulatory impact of TetC on its fusion partners. In this study, we have addressed this important issue. The protective 28-kDa glutathione S-transferase (GST) from Schistosoma haematobium (Sh28GST) was expressed either as a fusion to TetC or as the full-length Sh28GST alone in a nonvirulent aroA-attenuated strain of Salmonella enterica serovar Typhimurium. The Sh28GST proteins were soluble and stably expressed in Salmonella, as evaluated by Western blotting with TetC and/or Sh28GST antisera. Mice were immunized orally with a single dose of the live recombinant Salmonella. The constructs were stable in mice but, dramatically, only the strain expressing the TetC-Sh28GST fusion elicited significant antibody (Ab) responses directed against Sh28GST as determined by enzyme-linked immunosorbent assay. An analysis of the isotype profiles showed that these mice also produced anti-Sh28GST immunoglobulin A and GST-neutralizing assays revealed high levels of neutralizing Abs in sera. These are important correlates of protection in schistosomiasis. In addition, stimulation of spleen cells from immunized mice with Sh28GST Ag showed that both strains, expressing Sh28GST alone or the TetC-Sh28GST fusion, were able to stimulate the secretion of Th1-related cytokines (gamma interferon and interleukin 2) to comparable levels. Thus, TetC has modulated the immune responses generated against its fusion partner, Sh28GST, by markedly enhancing the Ab responses elicited. These results have important implications in the rational development of live vaccines.
Figures
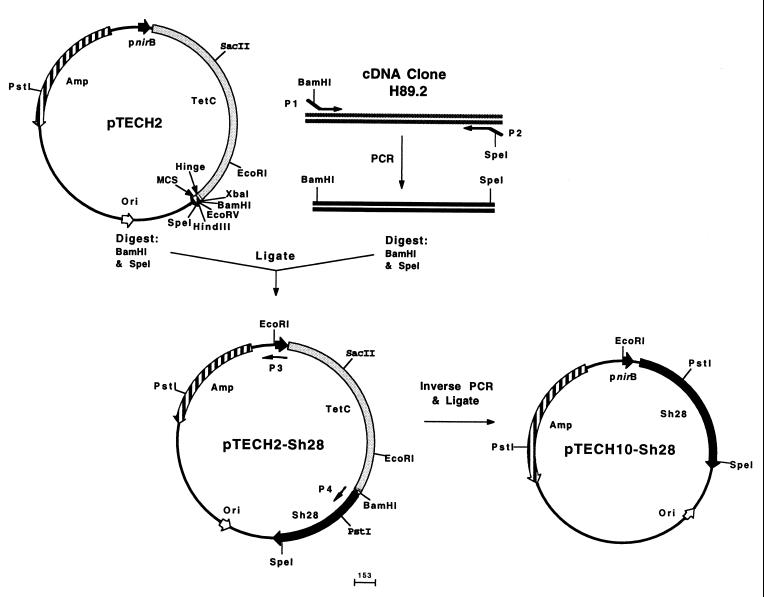
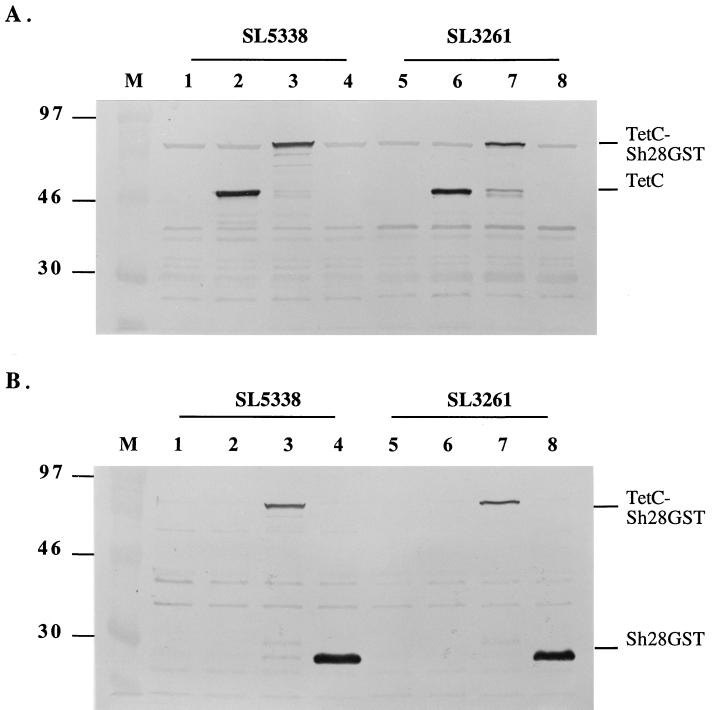
References
-
- Balloul J M, Grxych J M, Pierce R J, Capron A. A purified 28,000 dalton protein from Schistosoma mansoni adult worms protects rats and mice against experimental schistosomiasis. J Immunol. 1987;138:3448–3453. - PubMed
-
- Bergquist N R, Colley D G. Schistosomiasis vaccines: research to development. Parasitol Today. 1998;14:99–104. - PubMed
-
- Boulanger D, Reid G D, Sturrock R F, Wolowczuk I, Balloul J-M, Grezel D, Pierce R J, Otieno M F, Guerret S, Grimaud J A, Butterworth A E, Capron A. Immunization of mice and baboons with the recombinant Sm28GST affects both worm viability and fecundity after experimental infection with Schistosoma mansoni. Parasite Immunol. 1991;13:473–490. - PubMed
-
- Boulanger D, Warter A, Sellin B, Lindner V, Pierce R J, Chippaux J-P, Capron A. Vaccine potential of a recombinant glutathione S-transferase cloned from Schistosoma haematobium in primates experimentally infected with an homologous challenge. Vaccine. 1999;17:319–326. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
