

Targeted disruption of the class B scavenger receptor CD36 protects against atherosclerotic lesion development in mice
- PMID: 10772649
- PMCID: PMC300837
- DOI: 10.1172/JCI9259
Targeted disruption of the class B scavenger receptor CD36 protects against atherosclerotic lesion development in mice
Abstract
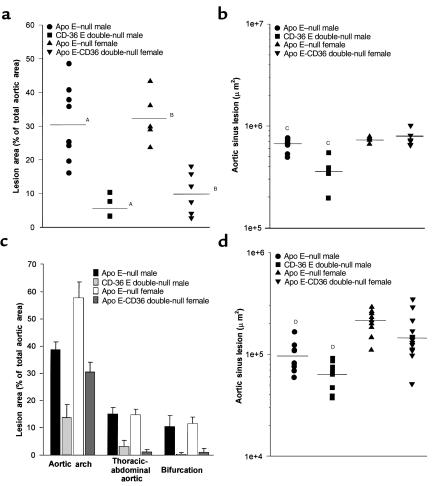
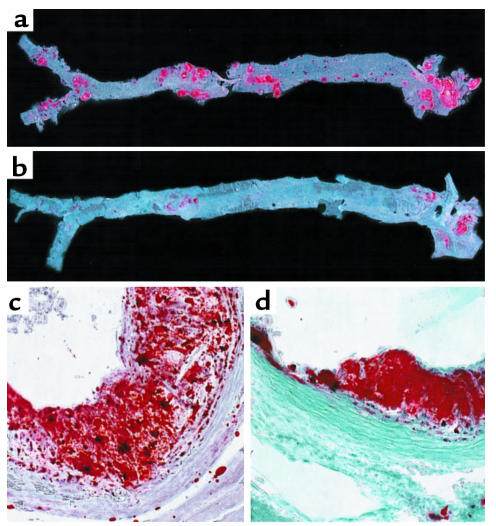
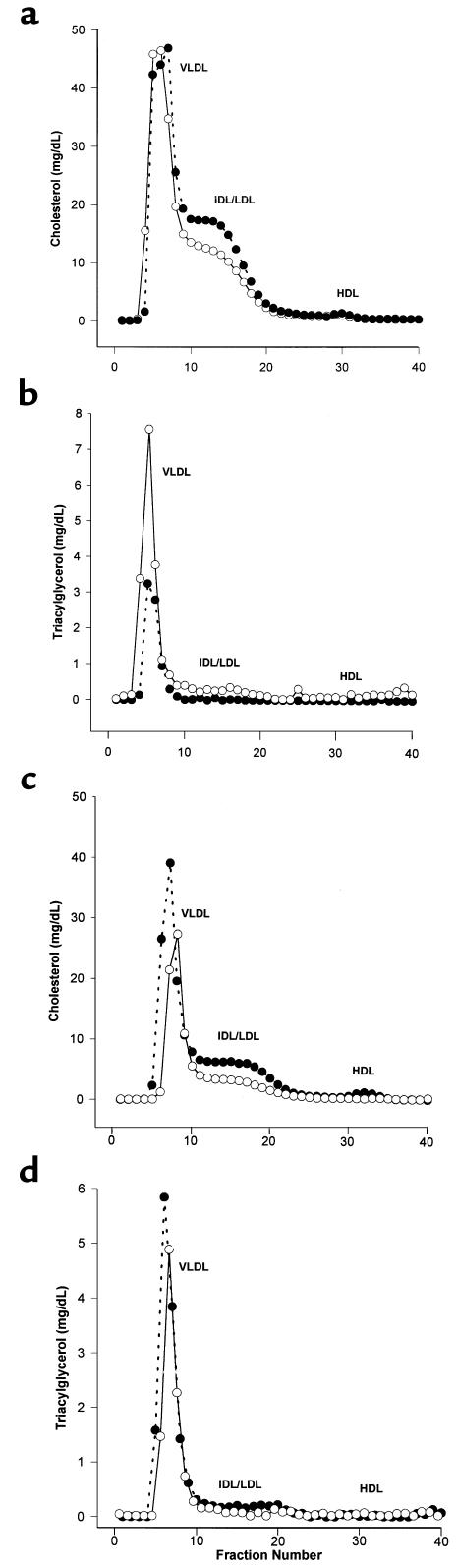
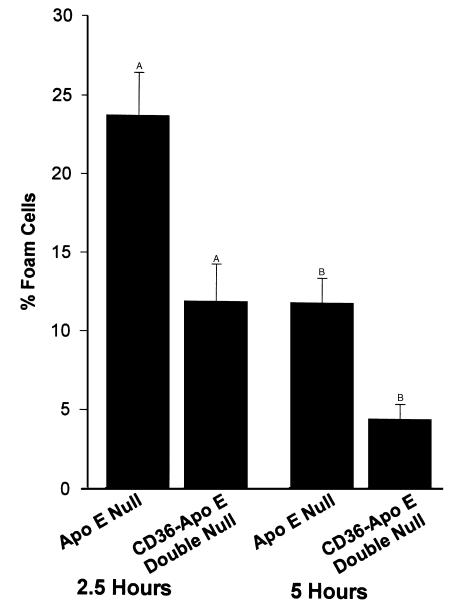
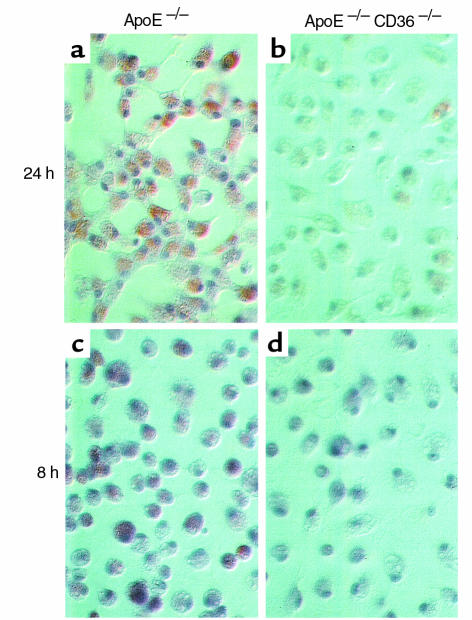
Macrophage scavenger receptors have been implicated as key players in the pathogenesis of atherosclerosis. To assess the role of the class B scavenger receptor CD36 in atherogenesis, we crossed a CD36-null strain with the atherogenic apo E-null strain and quantified lesion development. There was a 76.5% decrease in aortic tree lesion area (Western diet) and a 45% decrease in aortic sinus lesion area (normal chow) in the CD36-apo E double-null mice when compared with controls, despite alterations in lipoprotein profiles that often correlate with increased atherogenicity. Macrophages derived from CD36-apo E double-null mice bound and internalized more than 60% less copper-oxidized LDL and LDL modified by monocyte-generated reactive nitrogen species. A similar inhibition of in vitro lipid accumulation and foam cell formation after exposure to these ligands was seen. These results support a major role for CD36 in atherosclerotic lesion development in vivo and suggest that blockade of CD36 can be protective even in more extreme proatherogenic circumstances.
Figures

Comment in
-
Scavenging new insights into atherogenesis.J Clin Invest. 2000 Apr;105(8):1039-41. doi: 10.1172/JCI9919. J Clin Invest. 2000. PMID: 10772646 Free PMC article. No abstract available.
-
Peroxisome proliferator-activated receptor gamma ligands and atherosclerosis: ending the heartache.J Clin Invest. 2000 Sep;106(5):629-31. doi: 10.1172/JCI10909. J Clin Invest. 2000. PMID: 10974014 Free PMC article. No abstract available.
References
-
- Ross R. The pathogenesis of atherosclerosis: a perspective for the 1990s. Nature. 1993;362:801–809. - PubMed
-
- Marx N, Sukhova G, Murphy C, Libby P, Plutzky J. Macrophages in human atheroma contain PPAR-gamma: differentiation-dependent peroxisomal proliferator-activated receptor gamma (PPAR-gamma) expression and reduction of MMP-9 activity through PPAR-gamma activation in mononuclear phagocytes in vitro. Am J Pathol. 1998;153:17–23. - PMC - PubMed
-
- Boring L, Gosling J, Cleary M, Charo IF. Decreased lesion formation in CCR2–/– mice reveals a role for chemokines in the initiation of atherosclerosis. Nature. 1998;394:894–897. - PubMed
-
- de Villiers WJ, et al. Macrophage phenotype in mice deficient in both macrophage-colony-stimulating factor (op) and apolipoprotein E. Arterioscler Thromb Vasc Biol. 1998;18:631–640. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
