

Structure and mechanism of the aberrant ba(3)-cytochrome c oxidase from thermus thermophilus
- PMID: 10775261
- PMCID: PMC302014
- DOI: 10.1093/emboj/19.8.1766
Structure and mechanism of the aberrant ba(3)-cytochrome c oxidase from thermus thermophilus
Abstract
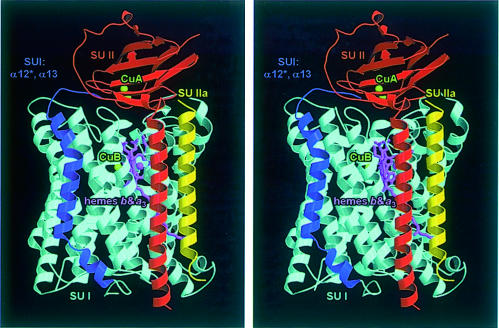
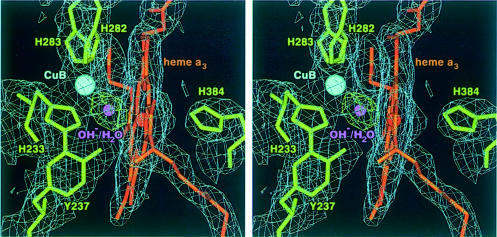
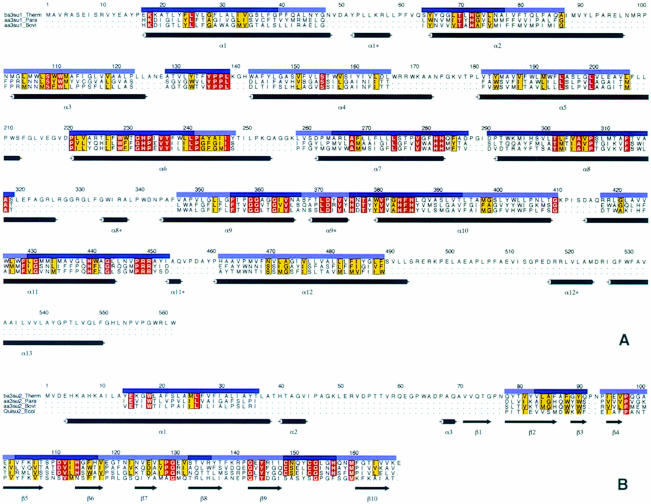
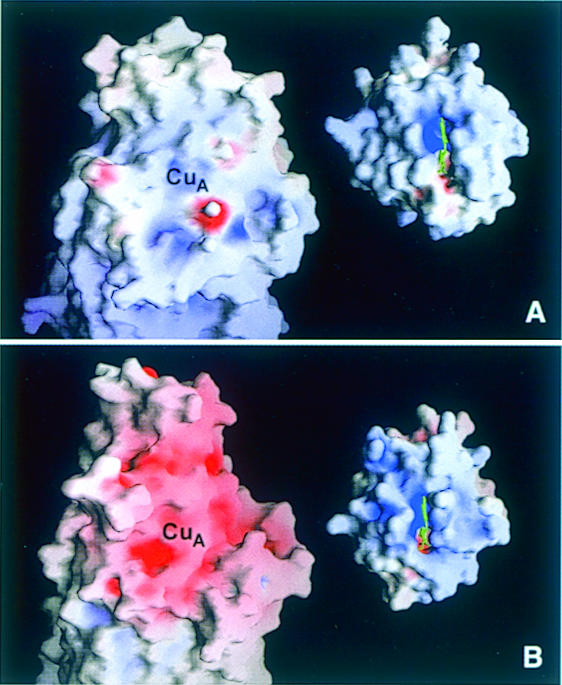
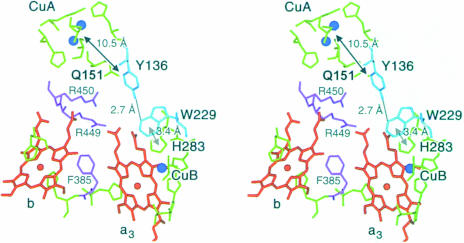
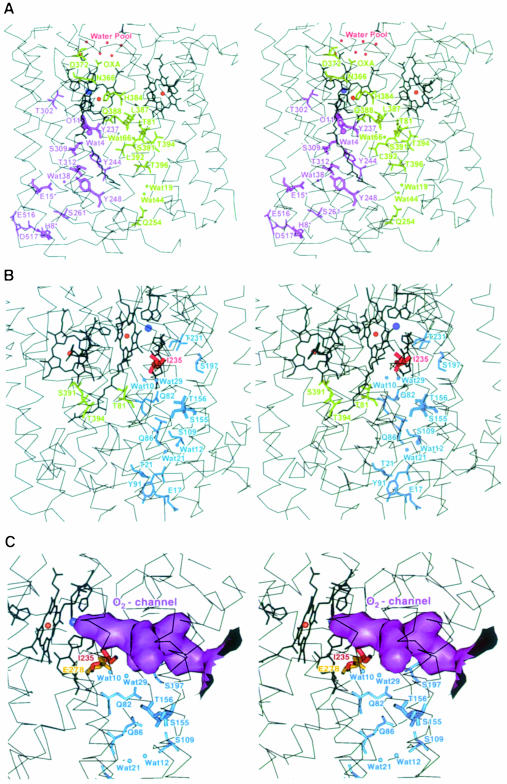
Cytochrome c oxidase is a respiratory enzyme catalysing the energy-conserving reduction of molecular oxygen to water. The crystal structure of the ba(3)-cytochrome c oxidase from Thermus thermophilus has been determined to 2.4 A resolution using multiple anomalous dispersion (MAD) phasing and led to the discovery of a novel subunit IIa. A structure-based sequence alignment of this phylogenetically very distant oxidase with the other structurally known cytochrome oxidases leads to the identification of sequence motifs and residues that seem to be indispensable for the function of the haem copper oxidases, e.g. a new electron transfer pathway leading directly from Cu(A) to Cu(B). Specific features of the ba(3)-oxidase include an extended oxygen input channel, which leads directly to the active site, the presence of only one oxygen atom (O(2-), OH(-) or H(2)O) as bridging ligand at the active site and the mainly hydrophobic character of the interactions that stabilize the electron transfer complex between this oxidase and its substrate cytochrome c. New aspects of the proton pumping mechanism could be identified.
Figures
References
-
- Adams M.W.W. and Kelly,R.M. (1998) Finding and using hyperthermophilic enzymes. Trends Biotechnol., 16, 329–332. - PubMed
-
- Barton G.J. (1993) ALSCRIPT: a tool to format multiple sequence alignments. Protein Eng., 6, 37–40. - PubMed
-
- Brünger A.T. et al. (1998) Crystallography and NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D, 54, 905–921. - PubMed
-
- Bushnell G.W., Louie,G.V. and Brayer,G.D. (1990) High-resolution three-dimensional structure of horse heart cytochrome c. J. Mol. Biol., 214, 585–595. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
