

The human immunodeficiency virus type 1 gp120 V2 domain mediates gp41-independent intersubunit contacts
- PMID: 10775580
- PMCID: PMC111965
- DOI: 10.1128/jvi.74.10.4448-4455.2000
The human immunodeficiency virus type 1 gp120 V2 domain mediates gp41-independent intersubunit contacts
Abstract
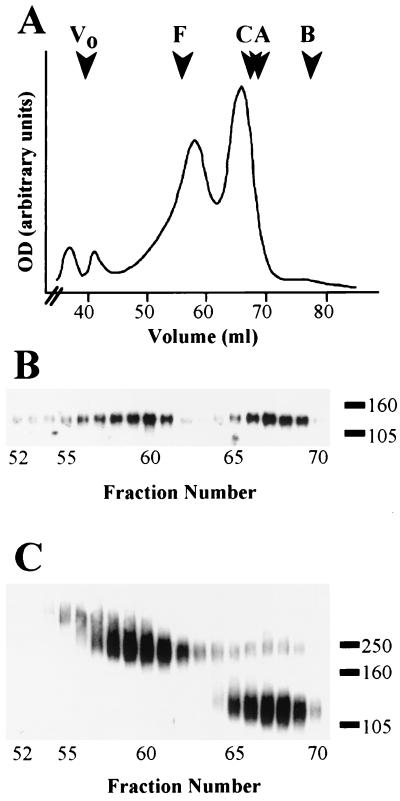
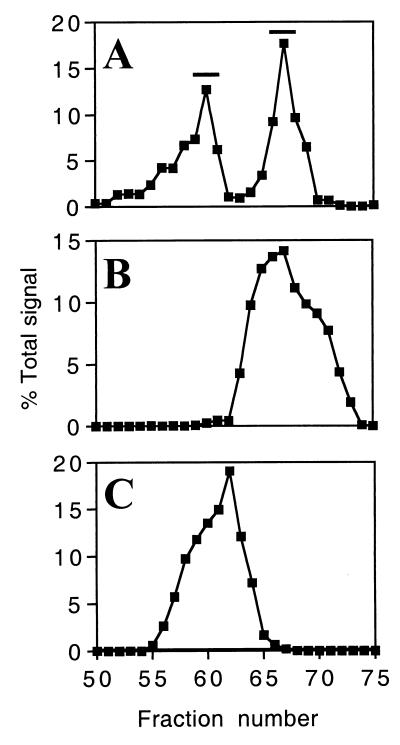
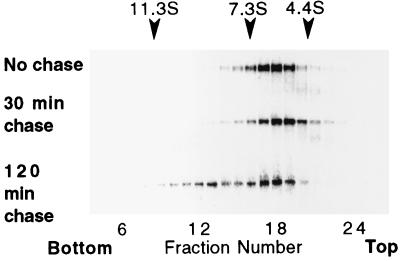
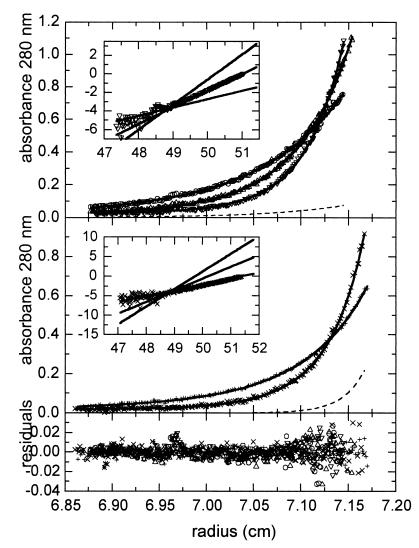
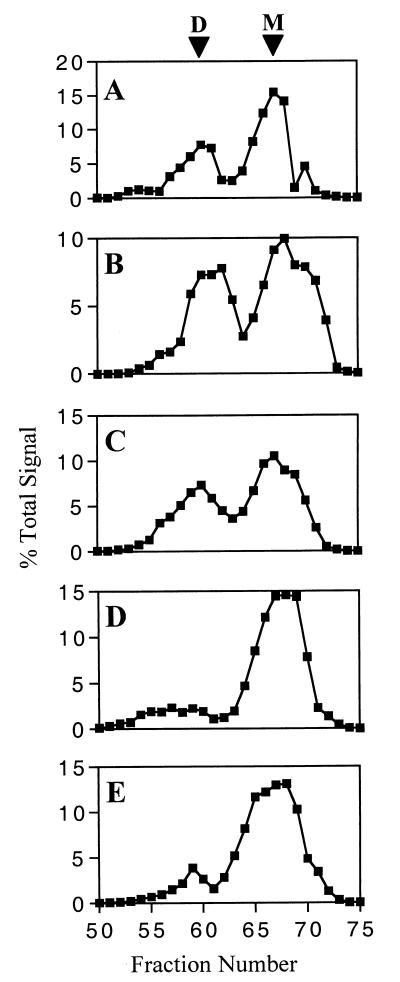
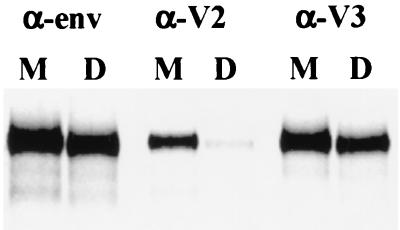
The envelope protein of human immunodeficiency virus type 1 HIV-1 undergoes proteolytic cleavage in the Golgi complex to produce subunits designated gp120 and gp41, which remain noncovalently associated. While gp41 has a well-characterized oligomeric structure, the maintenance of gp41-independent gp120 intersubunit contacts remains a contentious issue. Using recombinant vaccinia virus to achieve high-level expression of gp120 in mammalian cells combined with gel filtration analysis, we were able to isolate a discrete oligomeric form of gp120. Oligomerization of gp120 occurred intracellularly between 30 and 120 min after synthesis. Analysis by sedimentation equilibrium unequivocally identified the oligomeric species as a dimer. In order to identify the domains involved in the intersubunit contact, we expressed a series of gp120 proteins lacking various domains and assessed the effects of mutation on oligomeric structure. Deletion of the V1 or V3 loops had little effect on the relative amounts of monomer and dimer in comparison to wild-type gp120. In contrast, deletion of either all or part of the V2 loop drastically reduced dimer formation, indicating that this domain is required for intersubunit contact formation. Consistent with this, the V2 loop of the dimer was less accessible than that of the monomer to a specific monoclonal antibody. Previous studies have shown that while the V2 loop is not an absolute requirement for viral entry, the absence of this domain reduces viral resistance to neutralization by monoclonal antibodies or sera. We propose that the quaternary structure of gp120 may contribute to resistance to neutralization by limiting the exposure of conserved epitopes.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
