

L* protein of Theiler's murine encephalomyelitis virus is required for virus growth in a murine macrophage-like cell line
- PMID: 10775632
- PMCID: PMC112016
- DOI: 10.1128/jvi.74.10.4898-4901.2000
L* protein of Theiler's murine encephalomyelitis virus is required for virus growth in a murine macrophage-like cell line
Abstract
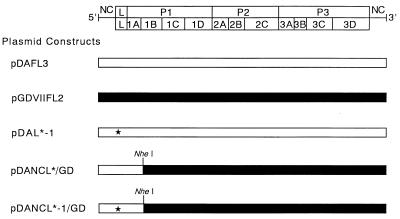
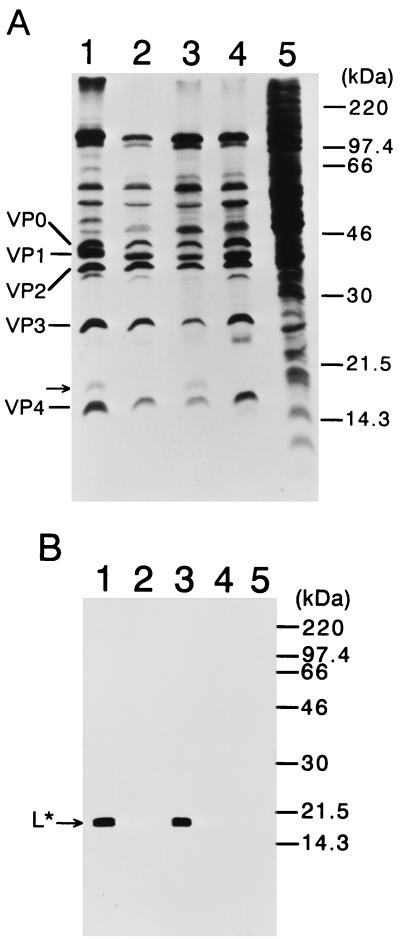
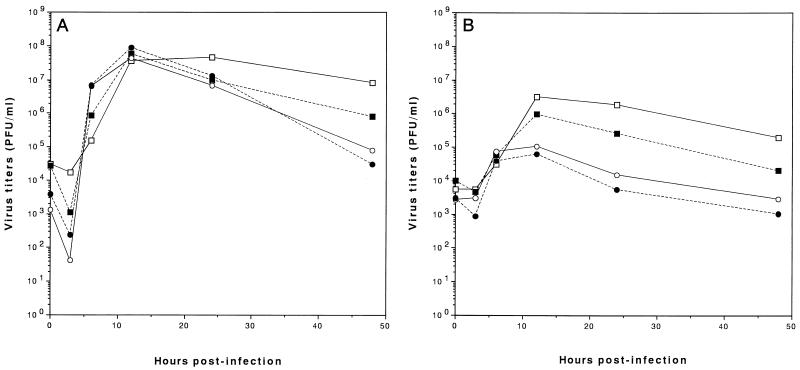
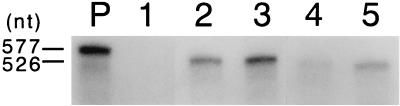
We sought to confirm the importance of L* protein for growth of Theiler's murine encephalomyelitis virus (TMEV) in a macrophage-like cell line, J774-1. The protein is out of frame with the polyprotein and synthesized in DA but not GDVII subgroup strains of TMEV. A recombinant virus, DANCL*/GD, which substitutes the DA 5' noncoding and L* coding regions for the corresponding regions of GDVII and synthesizes L* protein, grew with little restriction in J774-1 cells. In contrast, another recombinant virus, DANCL*-1/GD, which has an ACG rather than an AUG as the starting codon of L* protein at nucleotide 1079, resulting in no synthesis of L* protein, did not grow well. No significant difference between the rates of adsorption to J774-1 cells of these viruses was observed. RNase protection assay demonstrated that DANCL*/GD viral RNA significantly increased, whereas only a minimal increase was observed for DANCL*-1/GD. The present study suggests that L* protein is required for virus growth in macrophages.
Figures
Similar articles
-
Leader (L) of Theiler's murine encephalomyelitis virus (TMEV) is required for virus growth in a murine macrophage-like cell line.Virus Res. 2010 Feb;147(2):224-30. doi: 10.1016/j.virusres.2009.11.004. Epub 2009 Nov 20. Virus Res. 2010. PMID: 19932140
-
A lentiviral expression system demonstrates that L* protein of Theiler's murine encephalomyelitis virus (TMEV) is essential for virus growth in a murine macrophage-like cell line.Virus Res. 2005 Mar;108(1-2):23-8. doi: 10.1016/j.virusres.2004.07.009. Virus Res. 2005. PMID: 15681051
-
L* protein of the DA strain of Theiler's murine encephalomyelitis virus is important for virus growth in a murine macrophage-like cell line.J Virol. 1998 Jun;72(6):4950-5. doi: 10.1128/JVI.72.6.4950-4955.1998. J Virol. 1998. PMID: 9573263 Free PMC article.
-
Theiler's murine encephalomyelitis virus (TMEV): the role of a small out-of-frame protein in viral persistence and demyelination.Jpn J Infect Dis. 1999 Dec;52(6):228-33. Jpn J Infect Dis. 1999. PMID: 10738359 Review.
-
Theiler's virus persistence in the central nervous system of mice is associated with continuous viral replication and a difference in outcome of infection of infiltrating macrophages versus oligodendrocytes.Virus Res. 2005 Aug;111(2):214-23. doi: 10.1016/j.virusres.2005.04.010. Virus Res. 2005. PMID: 15893838 Review.
Cited by
-
Leader (L) and L* proteins of Theiler's murine encephalomyelitis virus (TMEV) and their regulation of the virus' biological activities.J Neuroinflammation. 2006 Aug 16;3:19. doi: 10.1186/1742-2094-3-19. J Neuroinflammation. 2006. PMID: 16911804 Free PMC article.
-
Theiler's virus L* protein is targeted to the mitochondrial outer membrane.J Virol. 2011 Apr;85(7):3690-4. doi: 10.1128/JVI.02023-10. Epub 2011 Jan 12. J Virol. 2011. PMID: 21228226 Free PMC article.
-
Transcriptome analysis following neurotropic virus infection reveals faulty innate immunity and delayed antigen presentation in mice susceptible to virus-induced demyelination.Brain Pathol. 2021 Nov;31(6):e13000. doi: 10.1111/bpa.13000. Epub 2021 Jul 6. Brain Pathol. 2021. PMID: 34231271 Free PMC article.
-
Evasion of antiviral innate immunity by Theiler's virus L* protein through direct inhibition of RNase L.PLoS Pathog. 2013;9(6):e1003474. doi: 10.1371/journal.ppat.1003474. Epub 2013 Jun 27. PLoS Pathog. 2013. PMID: 23825954 Free PMC article.
-
Infection of macrophage primary cultures by persistent and nonpersistent strains of Theiler's virus: role of capsid and noncapsid viral determinants.J Virol. 2004 Dec;78(23):13356-61. doi: 10.1128/JVI.78.23.13356-13361.2004. J Virol. 2004. PMID: 15542687 Free PMC article.
References
-
- Ahmed R, Morrison L A, Knipe D M. Persistence of viruses. In: Fields B N, Knipe D M, Howley P M, Chanock R M, Melnick J L, Monath T P, Roizman B, Straus S E, editors. Fields virology. 3rd ed. Vol. 1. Philadelphia, Pa: Lippincott-Raven; 1996. pp. 219–249.
-
- Chen H-H, Kong W-P, Zhang L, Ward P L, Roos R P. A picornaviral protein synthesized out of frame with the polyprotein plays a key role in a virus-induced immune-mediated demyelinating disease. Nat Med. 1995;1:927–931. - PubMed
-
- Clatch R J, Miller S D, Metzner R, Dal Canto M C, Lipton H L. Monocytes/macrophages isolated from the mouse central nervous system contain infectious Theiler's murine encephalomyelitis virus (TMEV) Virology. 1990;176:244–254. - PubMed
-
- Friedmann A, Lipton H L. Replication of Theiler's murine encephalomyelitis viruses in BHK21 cells: an electron microscopic study. Virology. 1980;101:389–398. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
