

Neurosteroid modulation of GABA IPSCs is phosphorylation dependent
- PMID: 10777770
- PMCID: PMC6773128
- DOI: 10.1523/JNEUROSCI.20-09-03067.2000
Neurosteroid modulation of GABA IPSCs is phosphorylation dependent
Abstract
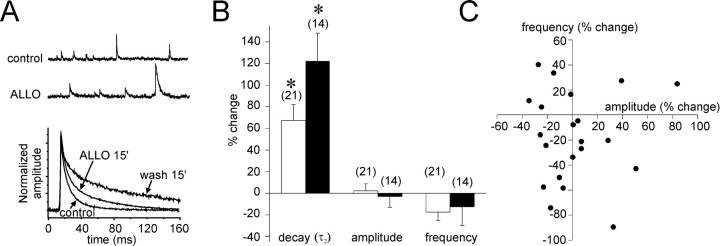
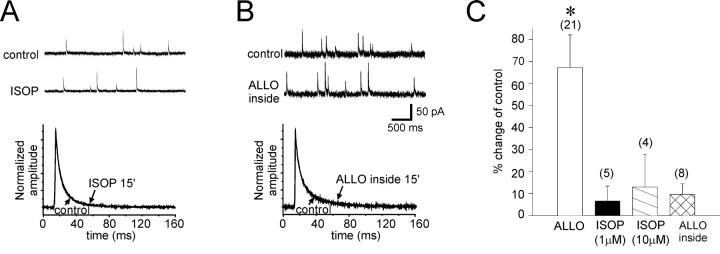
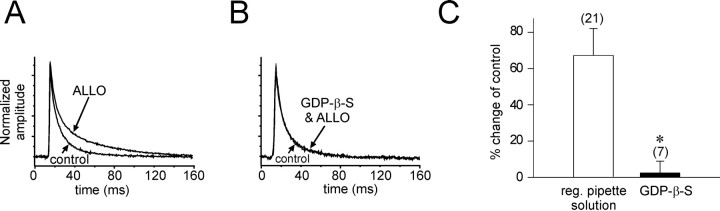
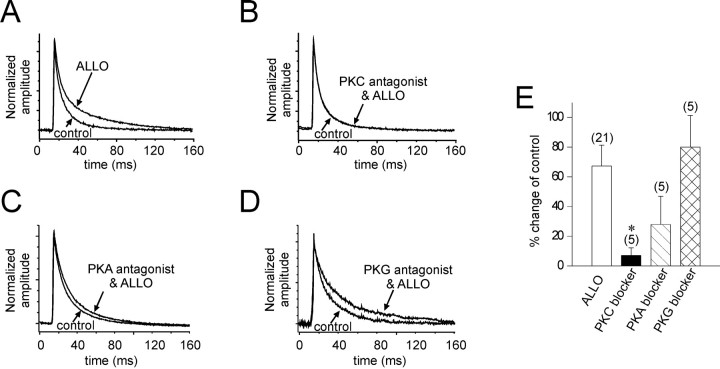
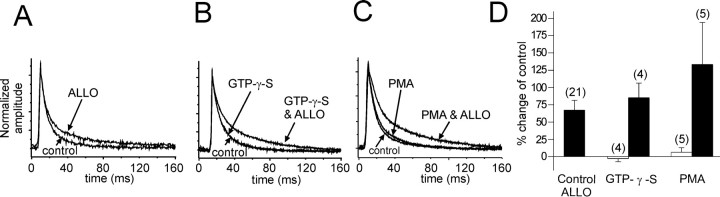
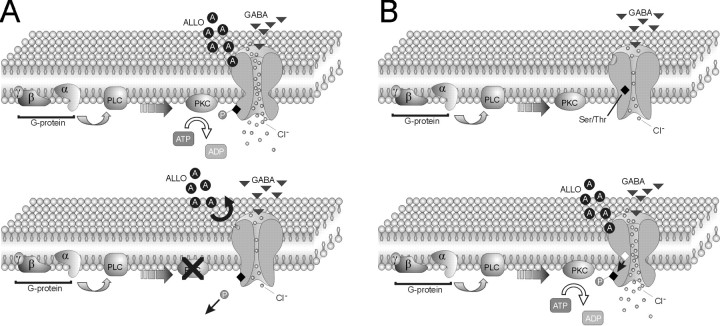
The neurosteroid 3alpha-hydroxy-5alpha-pregnan-20-one (allopregnanolone) facilitates GABA(A) receptor-mediated ionic currents via allosteric modulation of the GABA(A) receptor. Accordingly, allopregnanolone caused an increase in the slow decay time constant of spontaneous GABA-mediated IPSCs in magnocellular neurons recorded in hypothalamic slices. The allopregnanolone effect on IPSCs was inhibited by a G-protein antagonist as well as by blocking protein kinase C and, to a lesser extent, cAMP-dependent protein kinase activities. G-protein and protein kinase C activation in the absence of the neurosteroid had no effect on spontaneous IPSCs but enhanced the effect of subsequent allopregnanolone application. These findings together suggest that the neurosteroid modulation of GABA-mediated IPSCs requires G-protein and protein kinase activation, although not via a separate G-protein-coupled steroid receptor.
Figures
Similar articles
-
Neurosteroid modulation of synaptic and GABA-evoked currents in neurons from the rat medial preoptic nucleus.J Neurophysiol. 1999 Jul;82(1):143-51. doi: 10.1152/jn.1999.82.1.143. J Neurophysiol. 1999. PMID: 10400943
-
Modulation of GABAA receptor-mediated IPSCs by neuroactive steroids in a rat hypothalamo-hypophyseal coculture model.J Physiol. 1997 Apr 15;500 ( Pt 2)(Pt 2):475-85. doi: 10.1113/jphysiol.1997.sp022034. J Physiol. 1997. PMID: 9147331 Free PMC article.
-
Modulation of GABA(A) receptor function by neuroactive steroids: evidence for heterogeneity of steroid sensitivity of recombinant GABA(A) receptor isoforms.Can J Physiol Pharmacol. 1998 Sep;76(9):909-20. doi: 10.1139/cjpp-76-9-909. Can J Physiol Pharmacol. 1998. PMID: 10066142
-
Neurosteroid biosynthesis regulates sexually dimorphic fear and aggressive behavior in mice.Neurochem Res. 2008 Oct;33(10):1990-2007. doi: 10.1007/s11064-008-9718-5. Epub 2008 May 13. Neurochem Res. 2008. PMID: 18473173 Review.
-
Conditional regulation of neurosteroid sensitivity of GABAA receptors.Ann N Y Acad Sci. 2003 Dec;1007:29-36. doi: 10.1196/annals.1286.003. Ann N Y Acad Sci. 2003. PMID: 14993037 Review.
Cited by
-
Regulation of GABAARs by phosphorylation.Adv Pharmacol. 2015;72:97-146. doi: 10.1016/bs.apha.2014.11.008. Epub 2014 Dec 19. Adv Pharmacol. 2015. PMID: 25600368 Free PMC article. Review.
-
GABAA receptor currents in the dorsal motor nucleus of the vagus in females: influence of ovarian cycle and 5α-reductase inhibition.J Neurophysiol. 2019 Nov 1;122(5):2130-2141. doi: 10.1152/jn.00039.2019. Epub 2019 Oct 9. J Neurophysiol. 2019. PMID: 31596653 Free PMC article.
-
Preventing Phosphorylation of the GABA A R β3 Subunit Compromises the Behavioral Effects of Neuroactive Steroids.Front Mol Neurosci. 2022 Mar 31;15:817996. doi: 10.3389/fnmol.2022.817996. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35431797 Free PMC article.
-
Protein Kinase C-Dependent Effects of Neurosteroids on Synaptic GABAA Receptor Inhibition Require the δ-Subunit.Front Physiol. 2021 Oct 25;12:742838. doi: 10.3389/fphys.2021.742838. eCollection 2021. Front Physiol. 2021. PMID: 34759836 Free PMC article.
-
Sex-steroid-dependent plasticity of brain-stem autonomic circuits.Am J Physiol Regul Integr Comp Physiol. 2020 Jul 1;319(1):R60-R68. doi: 10.1152/ajpregu.00357.2019. Epub 2020 Jun 3. Am J Physiol Regul Integr Comp Physiol. 2020. PMID: 32493037 Free PMC article. Review.
References
-
- Baulieu EE. Neurosteroids: a novel function of the brain. Psychoneuroendocrinology. 1998;23:963–987. - PubMed
-
- Benke D, Fritschy JM, Trzeciak A, Bannwarth W, Mohler H. Distribution, prevalence, and drug binding profile of gamma-aminobutyric acid type A receptor subtypes differing in the beta-subunit variant. J Biol Chem. 1994;269:27100–27107. - PubMed
-
- Bixo M, Andersson A, Winblad B, Purdy RH, Backstrom T. Progesterone, 5alpha-pregnane-3,20-dione and 3alpha-hydroxy-5alpha-pregnane-20-one in specific regions of the human female brain in different endocrine states. Brain Res. 1997;764:173–178. - PubMed
-
- Brussaard AB, Kits KS, Baker RE, Willems WP, Leyting-Vermeulen JW, Voorn P, Smit AB, Bicknell RJ, Herbison AE. Plasticity in fast synaptic inhibition of adult oxytocin neurons caused by switch in GABA(A) receptor subunit expression. Neuron. 1997;19:1103–1114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources