

c-Raf regulates cell survival and retinal ganglion cell morphogenesis during neurogenesis
- PMID: 10777790
- PMCID: PMC6773115
- DOI: 10.1523/JNEUROSCI.20-09-03254.2000
c-Raf regulates cell survival and retinal ganglion cell morphogenesis during neurogenesis
Abstract
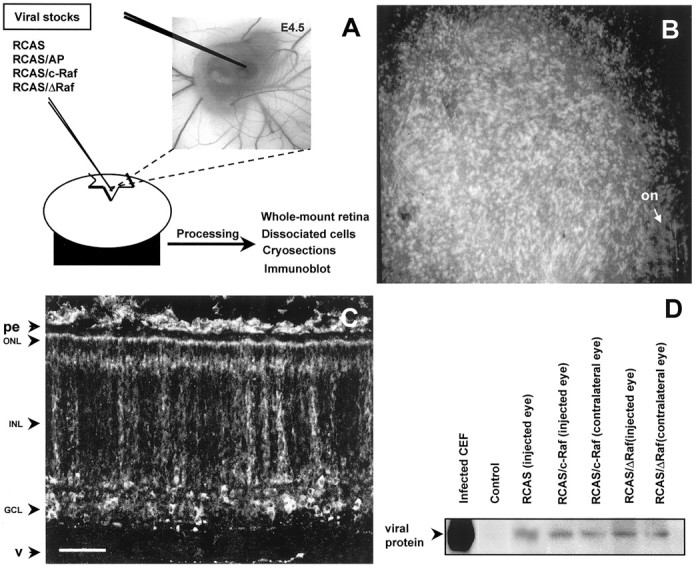
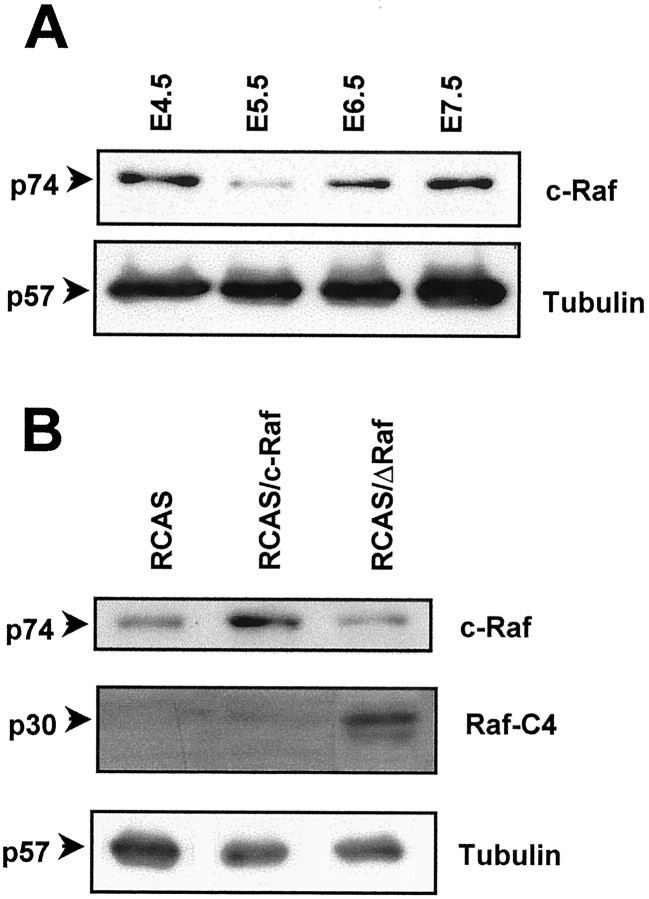
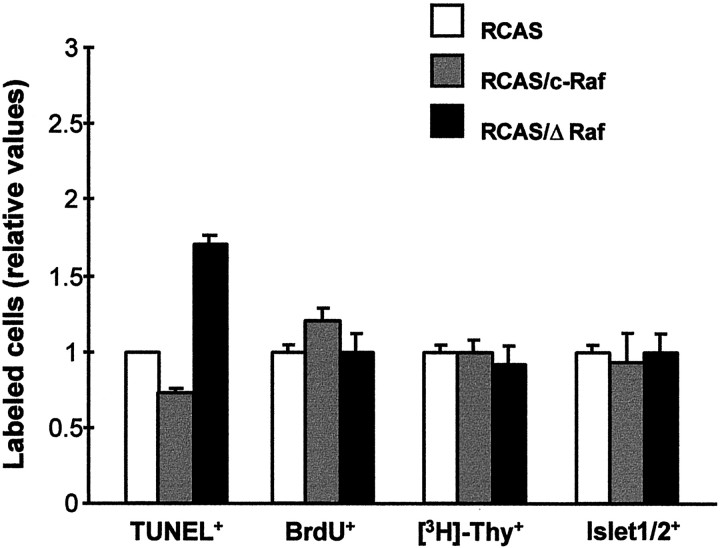
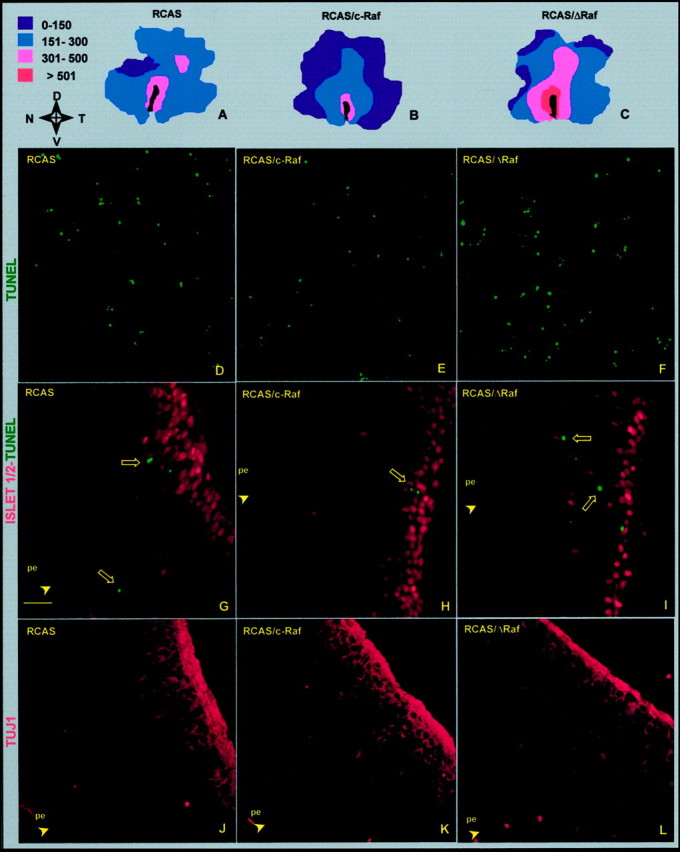
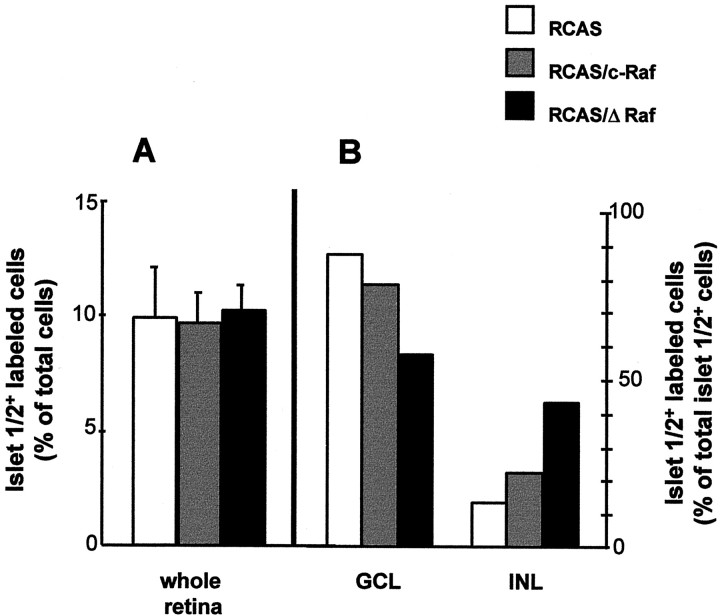
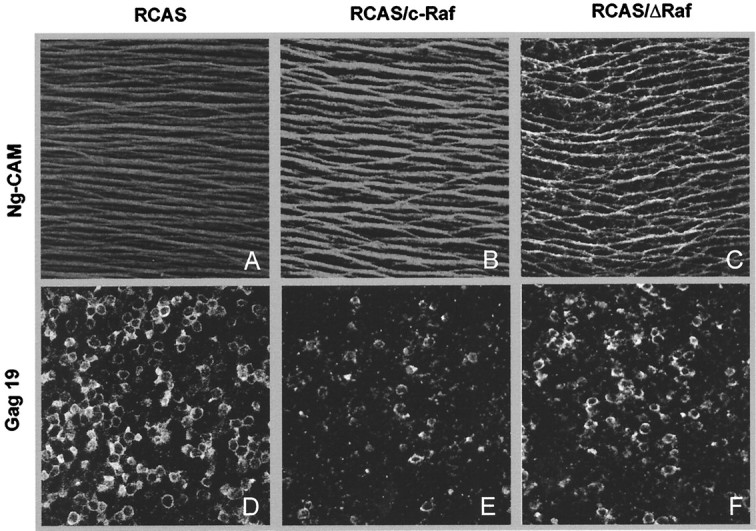
The signaling cascade Ras/Raf/mitogen-activated protein kinases modulates cell proliferation, differentiation, and survival, all key cellular processes during neural development. To better define the in vivo role of Raf during chick retinal neurogenesis, we interfered with Raf-dependent signaling during days 4.5 to 7.5 of embryonic development by expressing a dominant negative mutant of c-Raf (DeltaRaf), which blocks Ras-dependent Raf activation, and by overexpressing wild-type c-Raf. DeltaRaf expression induced an increase in cell death by apoptosis, whereas it did not affect overall cell proliferation and differentiation. In parallel, the number of Islet-1/2-positive and TUJ1-positive retinal ganglion cells were diminished in their definitive layer, whereas there was an increase in the number of mislocated Islet-1/2-positive cells. This disturbed morphogenesis correlated with a disruption of the optic fiber layer. Conversely, c-Raf overexpression caused moderate opposite effects on apoptosis. These results frame in vivo early neurogenesis processes in which c-Raf is essential.
Figures
References
-
- Alexiades MR, Cepko CL. Subsets of retinal progenitor display temporally regulated and distinct biases in the fates of their progeny. Development. 1997;124:1119–1131. - PubMed
-
- Altshuler DM, Turner DL, Cepko C. Specification of cell type in the vertebrate retina. In: Lau DM, Shatz CJ, editors. Development of the visual system. MIT; Cambridge, MA: 1991. pp. 37–58.
-
- Austin CP, Feldman DE, Ida JA, Cepko C. Vertebrate retinal ganglion cells are selected from competent progenitors by action of Notch. Development. 1995;121:3637–3650. - PubMed
-
- Barde Y-A. Trophic factors and neuronal survival. Neuron. 1989;2:1525–1534. - PubMed
-
- Blaschke AJ, Staley K, Chun J. Widespread programmed cell death in proliferative and postmitotic regions of the fetal cortex. Development. 1996;122:1165–1174. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous