

Ectopic expression of the neuropeptide pigment-dispersing factor alters behavioral rhythms in Drosophila melanogaster
- PMID: 10777797
- PMCID: PMC6773135
- DOI: 10.1523/JNEUROSCI.20-09-03339.2000
Ectopic expression of the neuropeptide pigment-dispersing factor alters behavioral rhythms in Drosophila melanogaster
Abstract
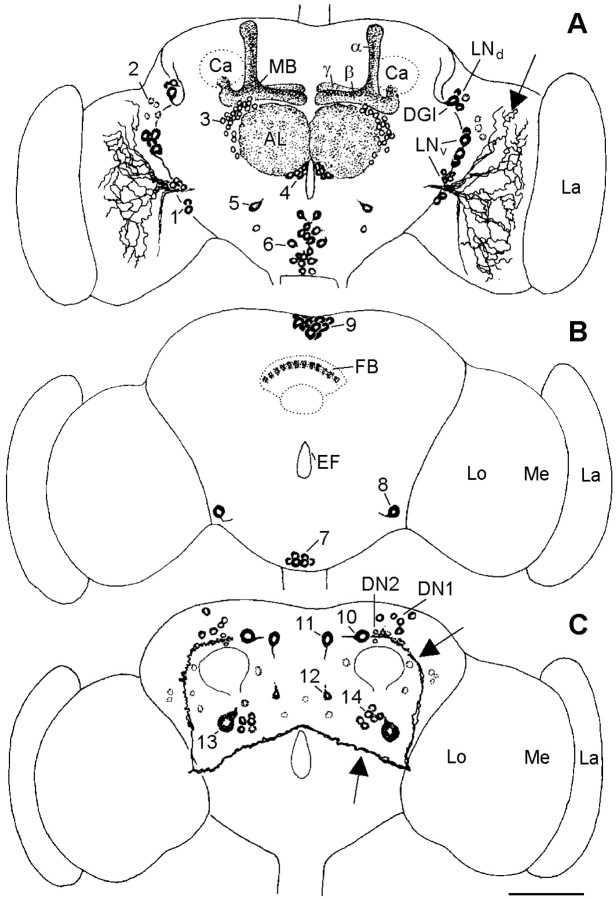
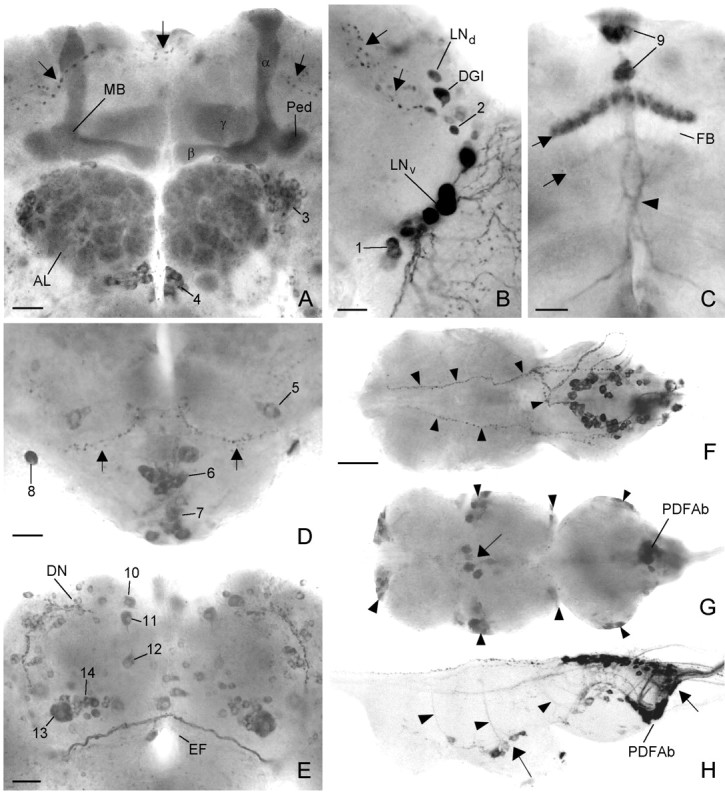
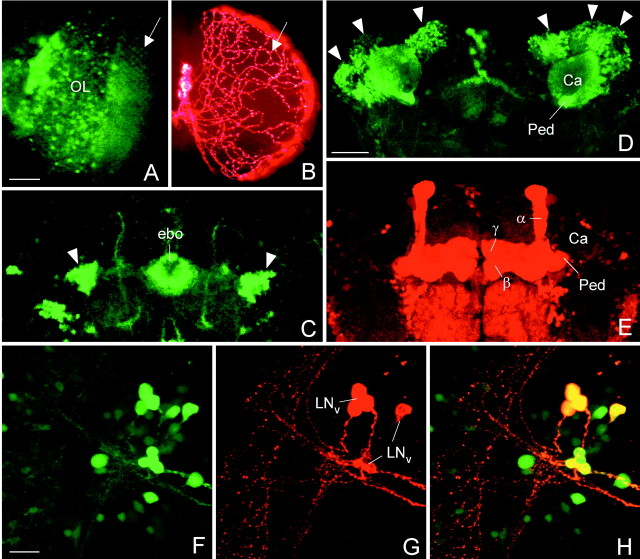
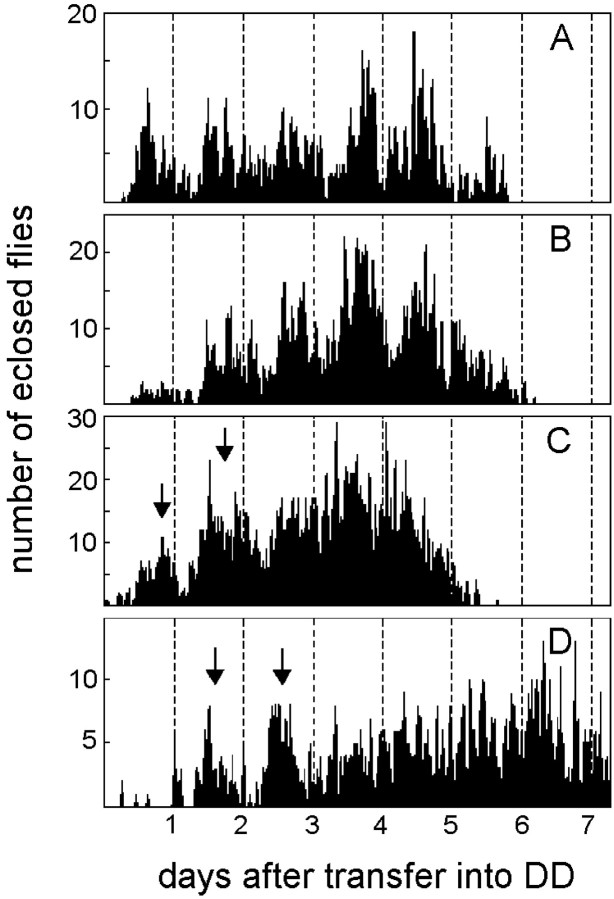
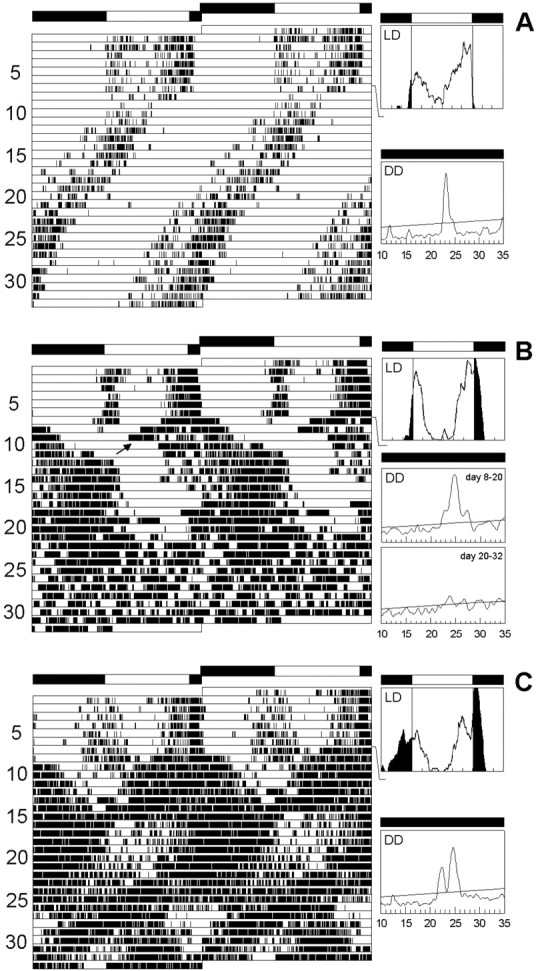
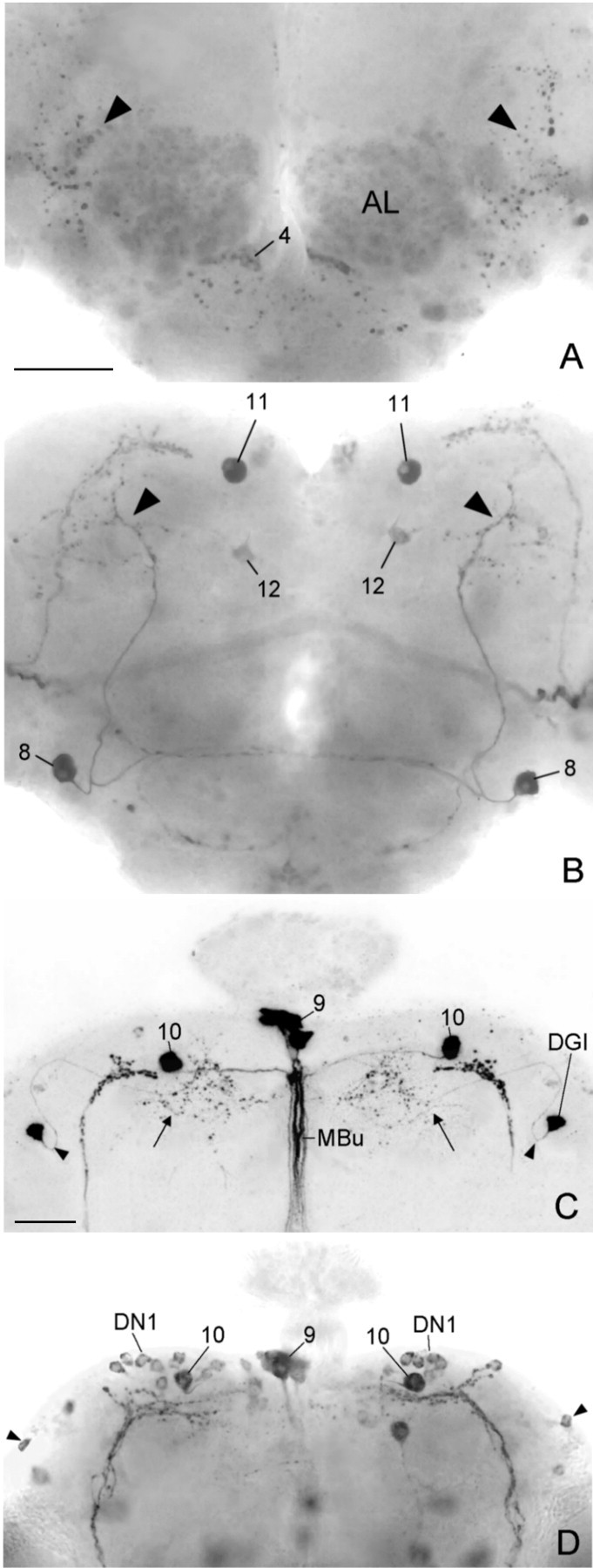
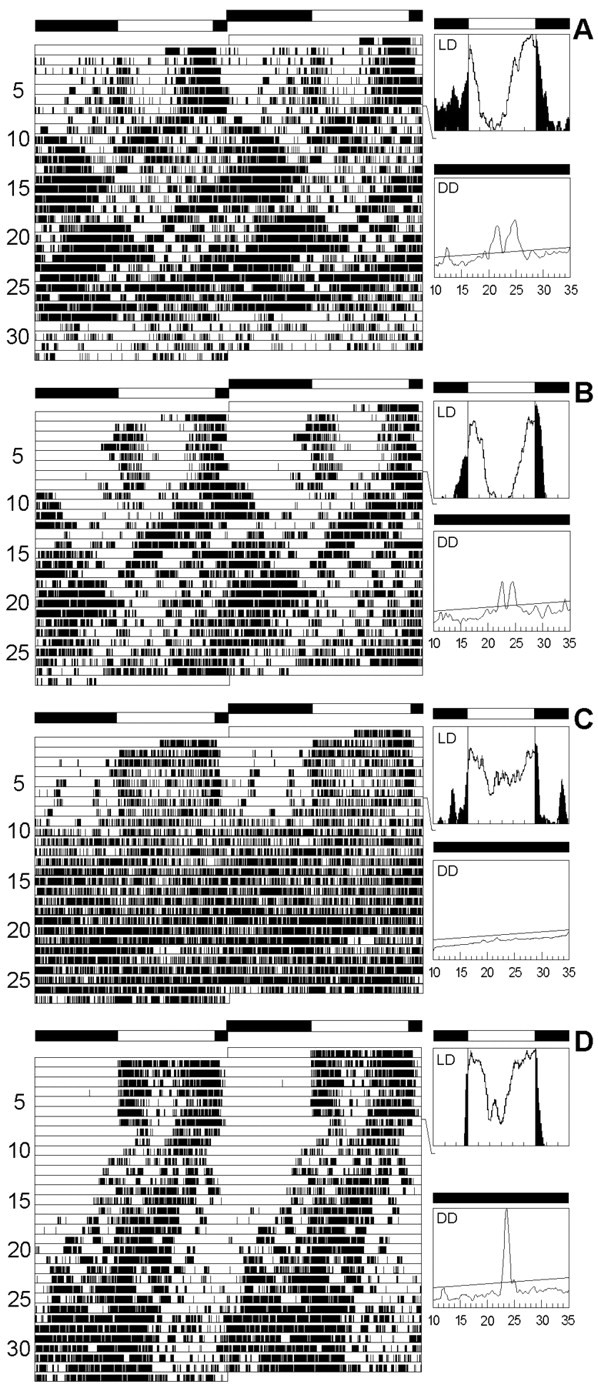
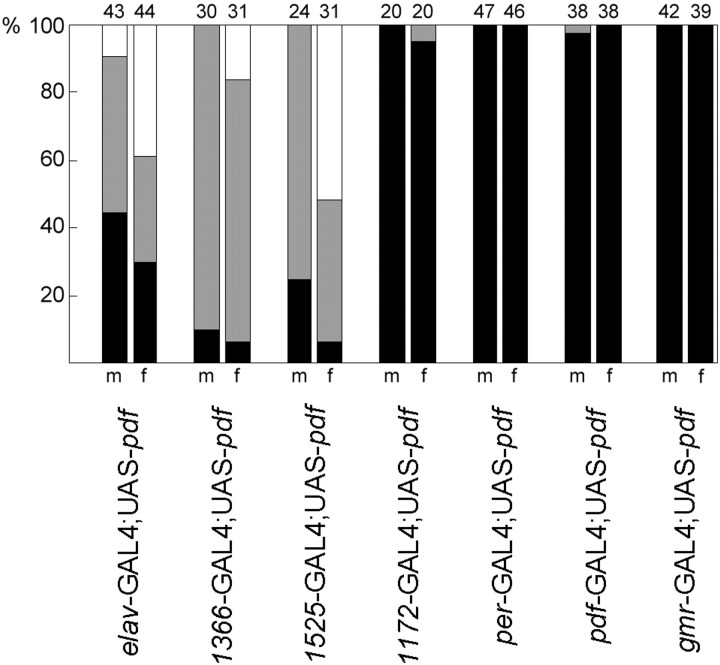
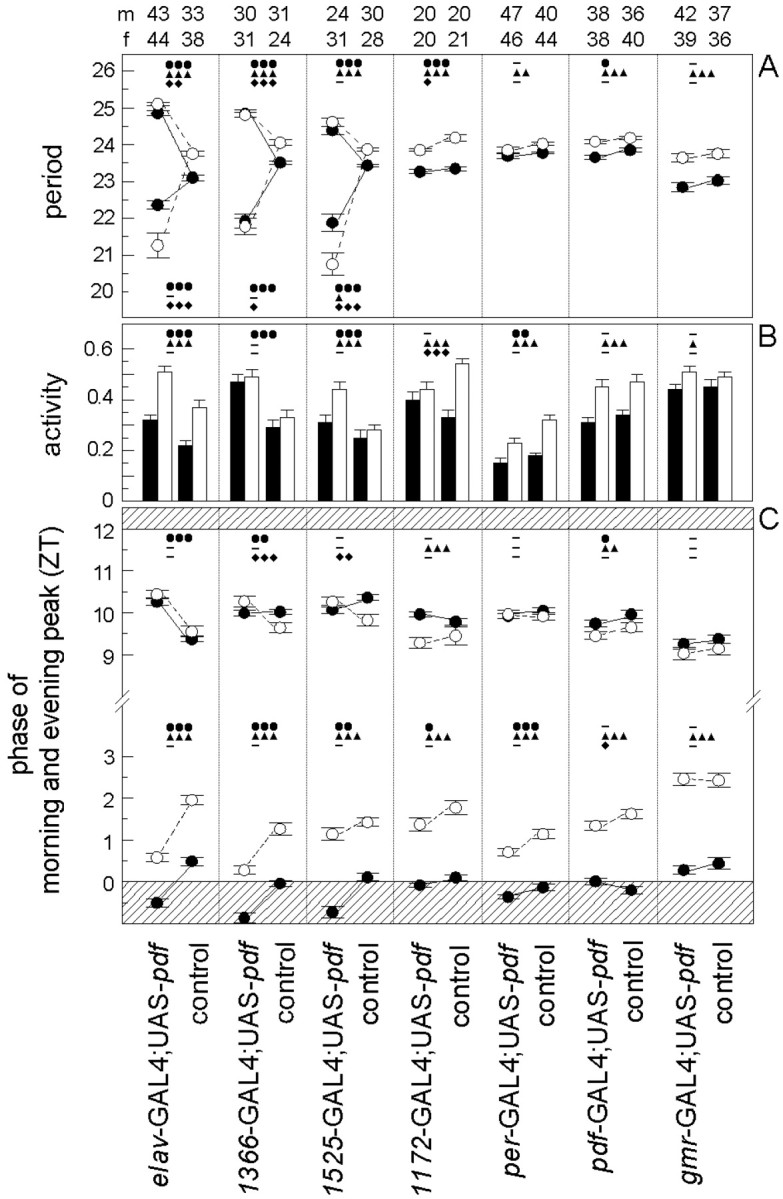
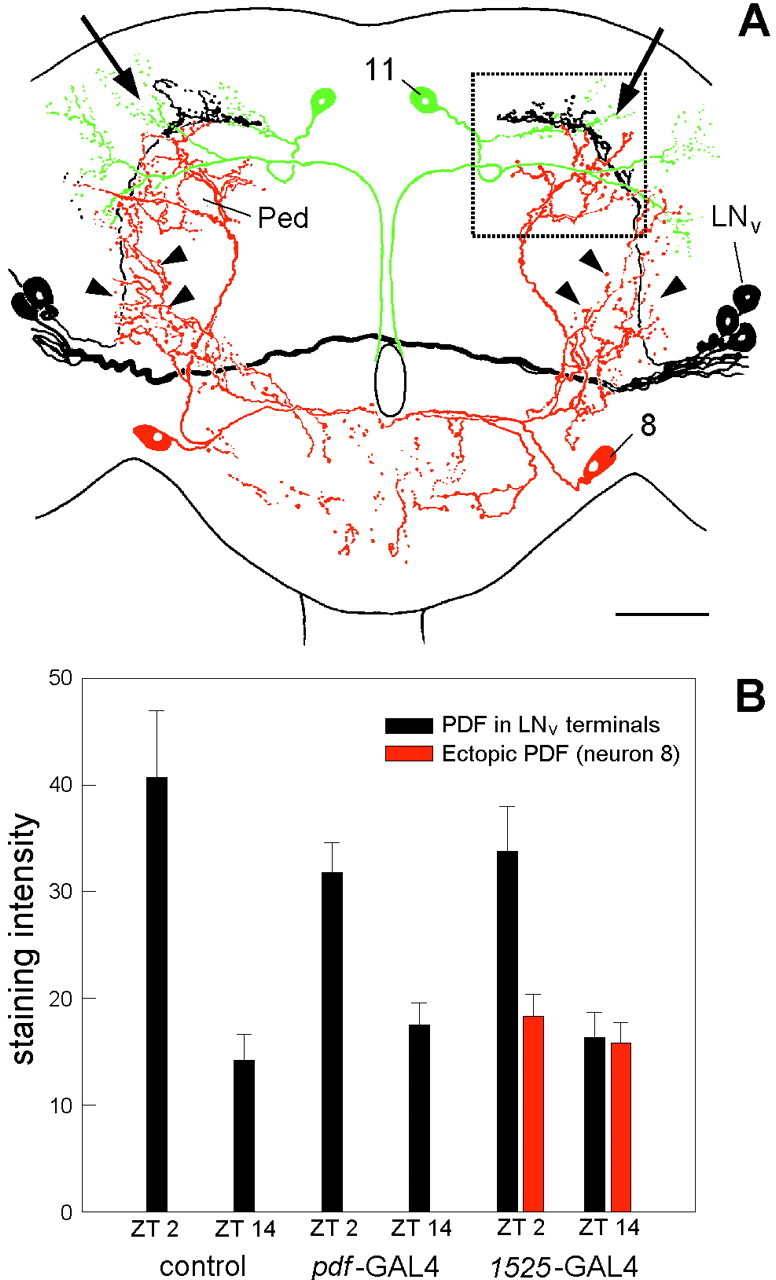
To study the function of the neuropeptide pigment-dispersing factor (PDF) in the circadian system of Drosophila, we misexpressed the pdf gene from the grasshopper Romalea in the CNS of Drosophila and investigated the effect of this on behavioral rhythmicity. pdf was either ectopically expressed in different numbers of neurons in the brain or the thoracical nervous system or overexpressed in the pacemaker neurons alone. We found severe alterations in the activity and eclosion rhythm of several but not all lines with ectopic pdf expression. Only ectopic pdf expression in neurons that projected into the dorsal central brain severely influenced activity rhythms. Therefore, we conclude that PDF acts as a neuromodulator in the dorsal central brain that is involved in the rhythmic control of behavior. Overexpression of pdf in the pacemaker neurons alone or in the other neurons that express the clock genes period (per) and timeless (tim) did not disturb the activity rhythm. Such flies still showed a rhythm in PDF accumulation in the central brain terminals. This rhythm was absent in the terminals of neurons that expressed PDF ectopically. Probably, PDF is rhythmically processed, transported, or secreted in neurons expressing per and tim, and additional PDF expression in these cells does not influence this rhythmic process. In neurons lacking per and tim, PDF appears to be continuously processed, leading to a constant PDF secretion at their nerve terminals. This may lead to conflicting signals in the rhythmic output pathway and result in a severely altered rhythmic behavior.
Figures
References
-
- Albers HE, Liou S-Y, Stopa EG, Zoeller RT. Neurotransmitter colocalization and circadian rhythms. Prog Brain Res. 1992;92:289–307. - PubMed
-
- Aréchiga H, Cortes JL, Farcia U, Rodrigues-Sosa L. Neuroendocrine correlates of circadian rhythmicity in Crustaceans. Am Zool. 1985;25:265–274.
-
- Brand A. GFP in Drosophila. Trends Genet. 1995;11:324–325. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases