

Extraretinal control of saccadic suppression
- PMID: 10777808
- PMCID: PMC6773104
- DOI: 10.1523/JNEUROSCI.20-09-03449.2000
Extraretinal control of saccadic suppression
Abstract
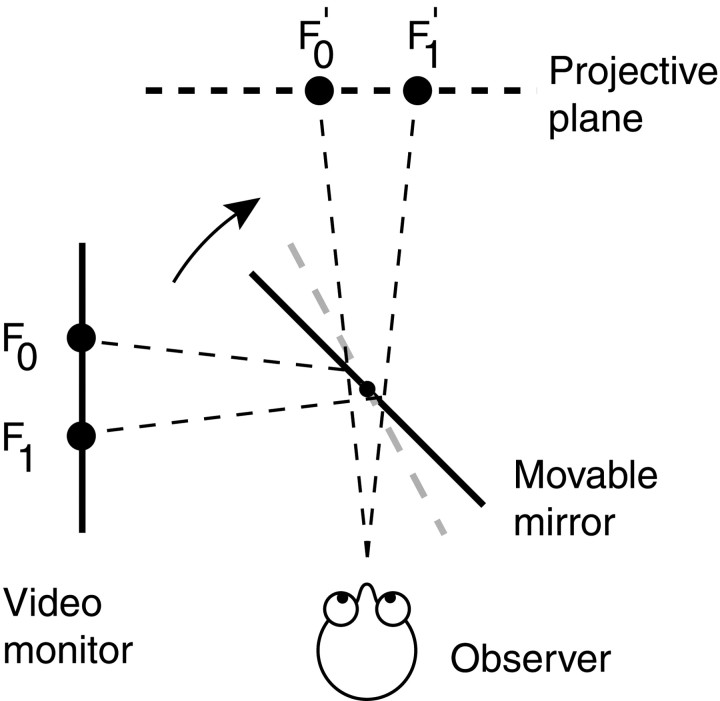
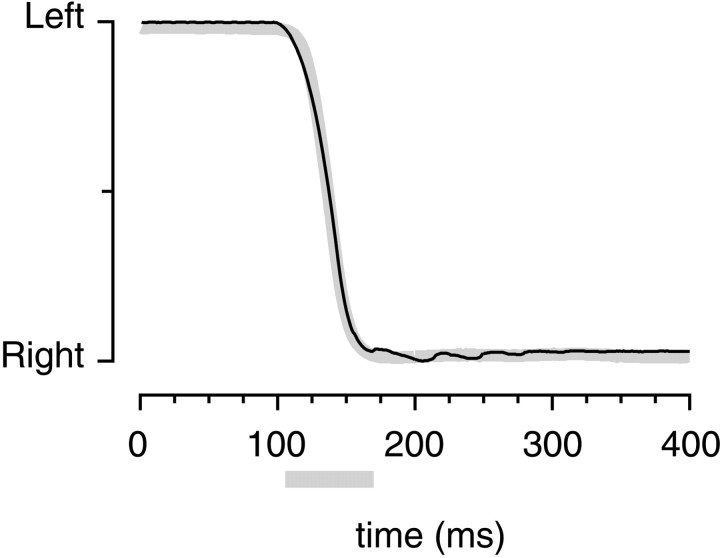
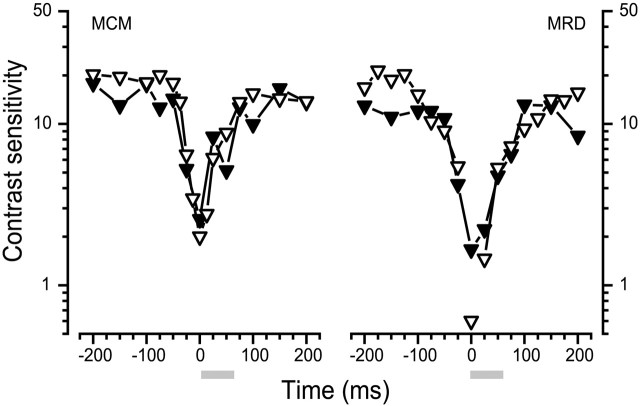
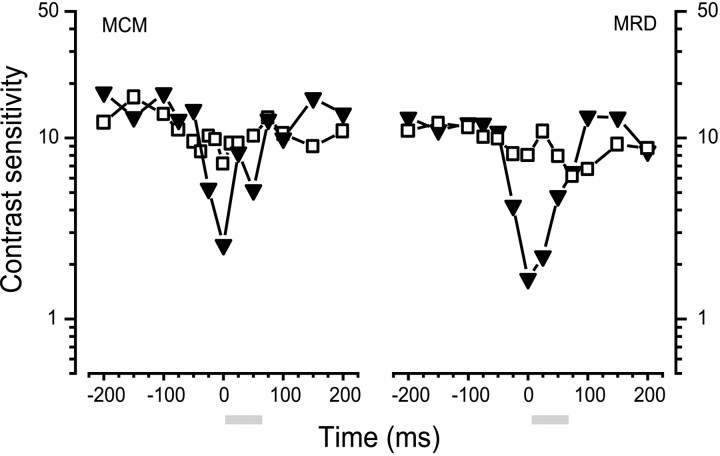
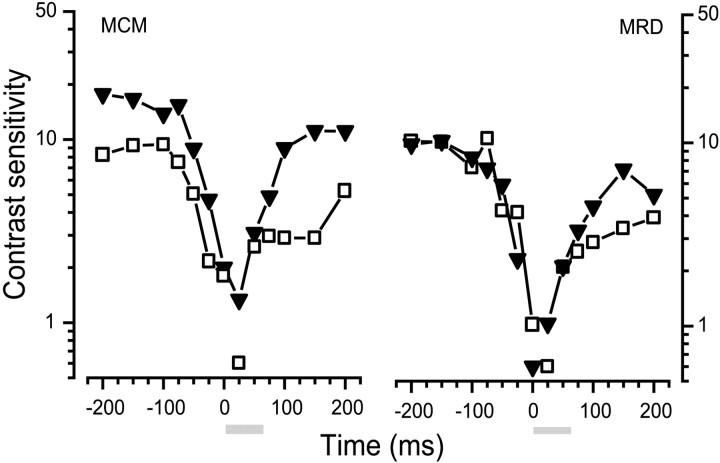
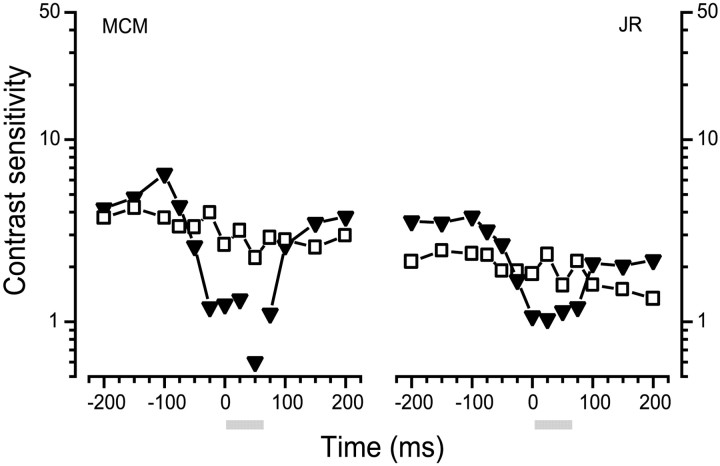
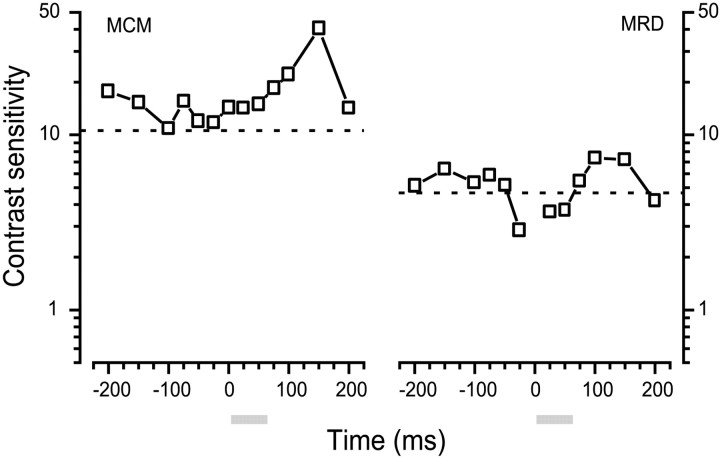
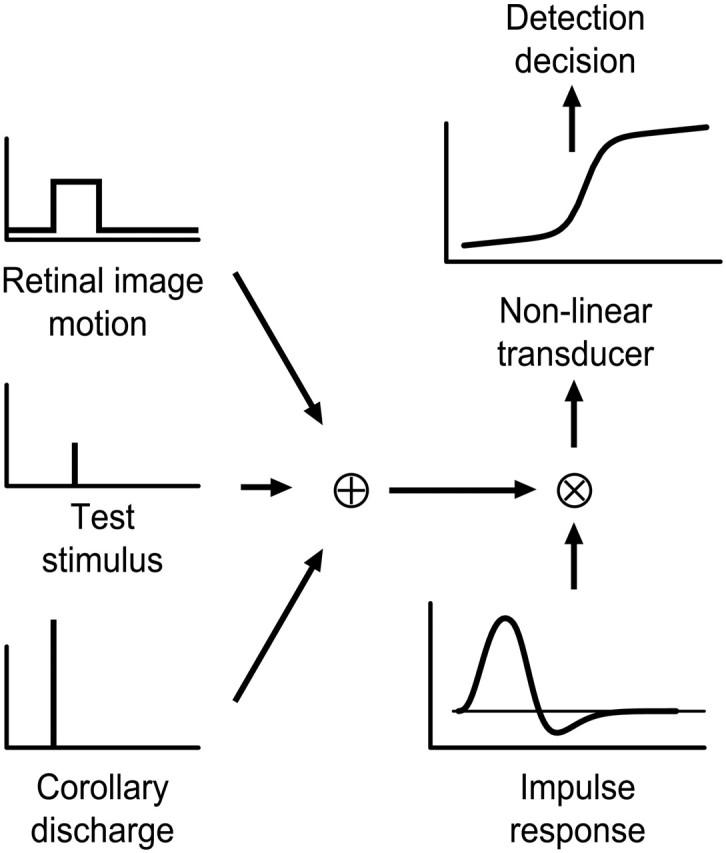
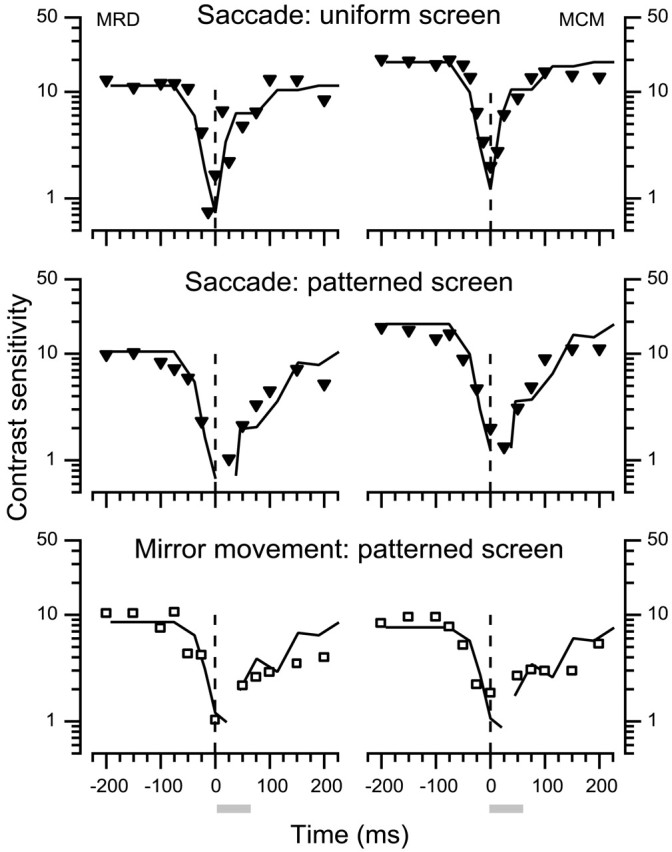
We measured the time course of saccadic suppression and tested whether suppression results entirely from retinal image motion or has an extraretinal source. We measured contrast thresholds for low-frequency gratings modulated either in luminance, at 17 cd/m(2) and 0.17 cd/m(2), or color at 17 cd/m(2). Gratings were flashed on a uniform background before, during, or after voluntary 12 degrees saccades and, additionally in the case of luminance modulated gratings, saccades simulated by mirror motion. A 10-fold decrease in contrast sensitivity was found for luminance-modulated gratings with saccades, but little suppression was found with simulated saccades. Adding high-contrast noise to the display increased the magnitude and the duration of the suppression during simulated saccades but had little effect on suppression produced by real saccades. Suppression anticipates saccades by 50 msec, is maximal at the moment of saccadic onset, and outlasts saccades by approximately 50 msec. At lower luminance, suppression is reduced, and its course is shallower than at higher luminance. Simulated saccades produce shallower suppression over a longer time course at both luminances. No suppression was found for chromatically modulated gratings. Differences between real and simulated saccades in the magnitude and time course of sensitivity loss suggest that saccadic suppression has an extraretinal component. We model the effects of saccades by adding a signal to the visual input, so as to saturate the nonlinear stage of visual processing and make detection of a test stimulus more difficult.
Figures
References
-
- Beeler GW. Visual threshold changes resulting from spontaneous saccadic eye movements. Vision Res. 1967;7:769–775. - PubMed
-
- Bridgeman B. Mechanisms of space constancy. In: Hein A, Jeannerod M, editors. Spatially oriented behavior. Springer; New York: 1983. pp. 263–279.
-
- Burr DC, Morrone MC. Impulse response functions for chromatic and achromatic stimuli. J Opt Soc Am A. 1993;10:1706–1713.
-
- Burr DC, Morrone MC. Temporal impulse response functions for luminance and color during saccades. Vision Res. 1996;36:2069–2078. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources