

Deficient gene expression in protein kinase inhibitor alpha Null mutant mice
- PMID: 10779334
- PMCID: PMC85637
- DOI: 10.1128/MCB.20.10.3442-3448.2000
Deficient gene expression in protein kinase inhibitor alpha Null mutant mice
Abstract
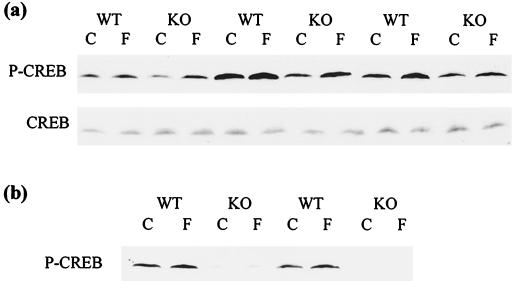
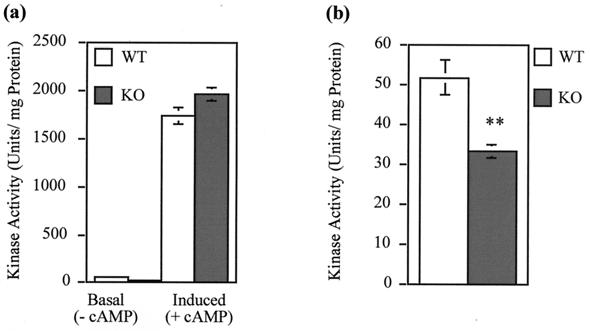
Protein kinase inhibitor (PKI) is a potent endogenous inhibitor of the cyclic AMP (cAMP)-dependent protein kinase (PKA). It functions by binding the free catalytic (C) subunit with a high affinity and is also known to export nuclear C subunit to the cytoplasm. The significance of these actions with respect to PKI's physiological role is not well understood. To address this, we have generated by homologous recombination mutant mice that are deficient in PKIalpha, one of the three isoforms of PKI. The mice completely lack PKI activity in skeletal muscle and, surprisingly, show decreased basal and isoproterenol-induced gene expression in muscle. Further examination revealed reduced levels of the phosphorylated (active) form of the transcription factor CREB (cAMP response element binding protein) in the knockouts. This phenomenon stems, at least in part, from lower basal PKA activity levels in the mutants, arising from a compensatory increase in the level of the RIalpha subunit of PKA. The deficit in gene induction, however, is not easily explained by current models of PKI function and suggests that PKI may play an as yet undescribed role in PKA signaling.
Figures

Similar articles
-
Cyclic AMP-dependent protein kinase regulates basal and cyclic AMP-stimulated but not phorbol ester-stimulated transcription of the tyrosine hydroxylase gene.J Neurochem. 1994 Sep;63(3):834-42. doi: 10.1046/j.1471-4159.1994.63030834.x. J Neurochem. 1994. PMID: 7914223
-
Cyclic AMP-mediated upregulation of the expression of neuronal NO synthase in human A673 neuroepithelioma cells results in a decrease in the level of bioactive NO production: analysis of the signaling mechanisms that are involved.Biochemistry. 2004 Jun 8;43(22):7197-206. doi: 10.1021/bi0302191. Biochemistry. 2004. PMID: 15170357
-
Characterization of PKIgamma, a novel isoform of the protein kinase inhibitor of cAMP-dependent protein kinase.J Biol Chem. 1997 Jul 18;272(29):18169-78. doi: 10.1074/jbc.272.29.18169. J Biol Chem. 1997. PMID: 9218452
-
Signal transduction through the cAMP-dependent protein kinase.Mol Cell Biochem. 1993 Nov;127-128:179-86. doi: 10.1007/BF01076769. Mol Cell Biochem. 1993. PMID: 7935349 Review.
-
Protein kinase inhibitor peptide (PKI): a family of endogenous neuropeptides that modulate neuronal cAMP-dependent protein kinase function.Neuropeptides. 2006 Feb;40(1):23-34. doi: 10.1016/j.npep.2005.10.002. Epub 2006 Jan 26. Neuropeptides. 2006. PMID: 16442618 Review.
Cited by
-
Modulation of polycystic kidney disease by G-protein coupled receptors and cyclic AMP signaling.Cell Signal. 2020 Aug;72:109649. doi: 10.1016/j.cellsig.2020.109649. Epub 2020 Apr 23. Cell Signal. 2020. PMID: 32335259 Free PMC article. Review.
-
PKA catalytic subunit compartmentation regulates contractile and hypertrophic responses to β-adrenergic signaling.J Mol Cell Cardiol. 2014 Jan;66:83-93. doi: 10.1016/j.yjmcc.2013.11.001. Epub 2013 Nov 10. J Mol Cell Cardiol. 2014. PMID: 24225179 Free PMC article.
-
Protein kinase A inhibitor proteins (PKIs) divert GPCR-Gαs-cAMP signaling toward EPAC and ERK activation and are involved in tumor growth.FASEB J. 2020 Oct;34(10):13900-13917. doi: 10.1096/fj.202001515R. Epub 2020 Aug 24. FASEB J. 2020. PMID: 32830375 Free PMC article.
-
Edmond Fischer's kinase legacy: History of the protein kinase inhibitor and protein kinase A.IUBMB Life. 2023 Apr;75(4):311-323. doi: 10.1002/iub.2714. Epub 2023 Feb 28. IUBMB Life. 2023. PMID: 36855225 Free PMC article.
-
MicroRNA-155 is required for Mycobacterium bovis BCG-mediated apoptosis of macrophages.Mol Cell Biol. 2012 Jun;32(12):2239-53. doi: 10.1128/MCB.06597-11. Epub 2012 Apr 2. Mol Cell Biol. 2012. PMID: 22473996 Free PMC article.
References
-
- Amieux P S, Cummings D E, Motamed K, Brandon E P, Wailes L A, Le K, Idzerda R L, McKnight G S. Compensatory regulation of RIalpha protein levels in protein kinase A mutant mice. J Biol Chem. 1997;272:3993–3998. - PubMed
-
- Baude E J, Dignam S S, Reimann E M, Uhler M D. Evidence for the importance of hydrophobic residues in the interactions between the cAMP-dependent protein kinase catalytic subunit and the protein kinase inhibitors. J Biol Chem. 1994;269:18128–18133. - PubMed
-
- Beale E G, Dedman J R, Means A R. Isolation and characterization of a protein from rat testis which inhibits cyclic AMP-dependent protein kinase and phosdiesterase. J Biol Chem. 1977;252:6322–6327. - PubMed
-
- Beebe S J, Koch S R, Chu D T, Corbin J D, Granner D K. Regulation of phosphoenolpyruvate carboxykinase gene transcription in H4IIE hepatoma cells: evidence for a primary role of the catalytic subunit of 3′,5′-cyclic adenosine monophosphate-dependent protein kinase. Mol Endocrinol. 1987;1:639–647. - PubMed
-
- Brandon E P, Gerhold K A, Qi M, McKnight G S, Idzerda R L. Derivation of novel embryonic stem cell lines and targeting of cyclic AMP-dependent protein kinase genes. Recent Prog Horm Res. 1995;50:403–408. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases