

UV-Induced stabilization of c-fos and other short-lived mRNAs
- PMID: 10779351
- PMCID: PMC85654
- DOI: 10.1128/MCB.20.10.3616-3625.2000
UV-Induced stabilization of c-fos and other short-lived mRNAs
Abstract
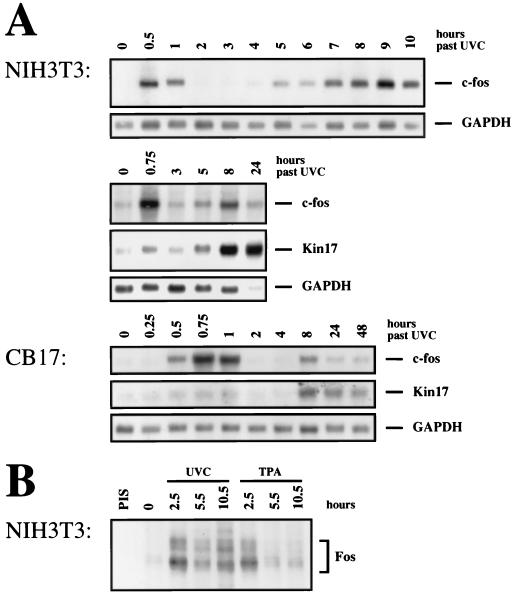
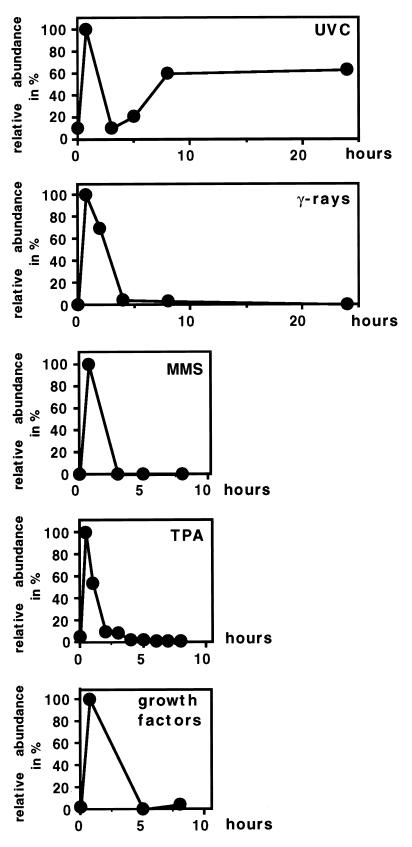
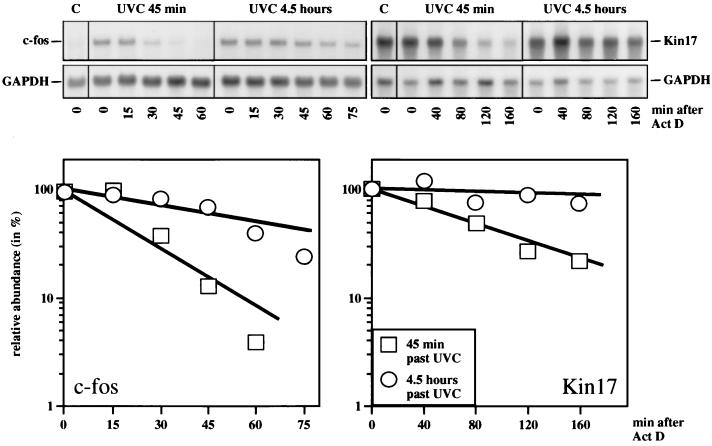
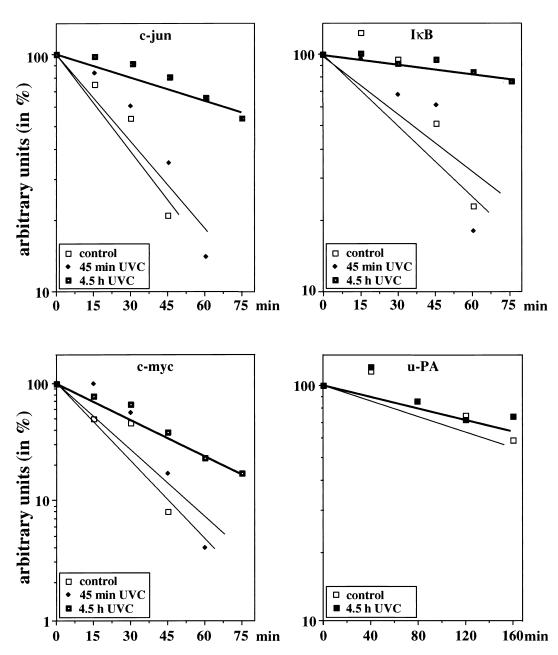
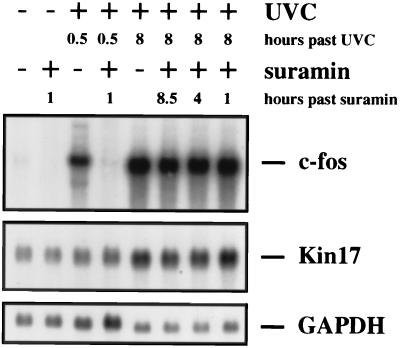
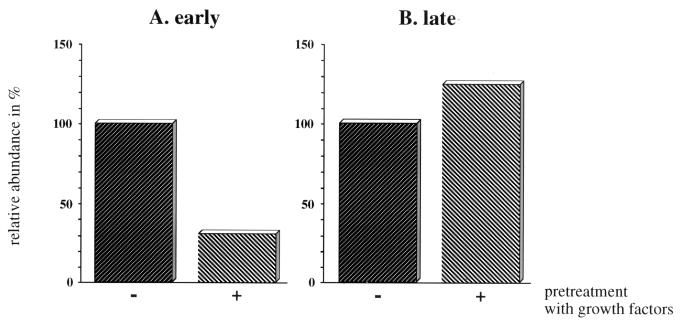
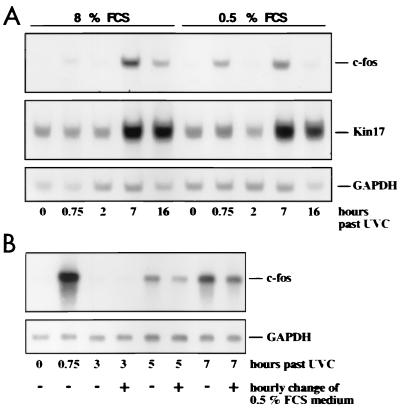
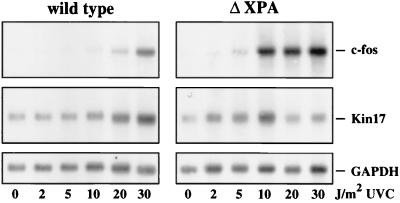
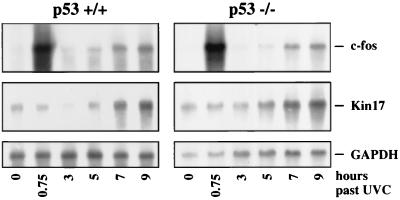
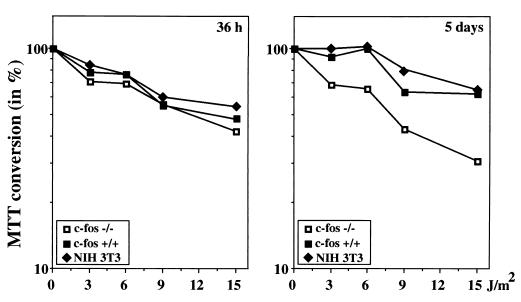
Irradiation of cells with short-wavelength ultraviolet light (UVC) changes the program of gene expression, in part within less than 15 min. As one of the immediate-early genes in response to UV, expression of the oncogene c-fos is upregulated. This immediate induction is regulated at the transcriptional level and is transient in character, due to the autocatalyzed shutoff of transcription and the rapid turnover of c-fos mRNA. In an experiment analyzing the kinetics of c-fos mRNA expression in murine fibroblasts irradiated with UVC, we found that, in addition to the initial transient induction, c-fos mRNA accumulated in a second wave starting at 4 to 5 h after irradiation, reaching a maximum at 8 h, and persisting for several more hours. It was accompanied by an increase in Fos protein synthesis. The second peak of c-fos RNA was caused by an UV dose-dependent increase in mRNA half-life from about 10 to 60 min. With similar kinetics, the mRNAs of other UV target genes (i.e., the Kin17 gene, c-jun, IkappaB, and c-myc) were stabilized (e.g., Kin17 RNA from 80 min to more than 8 h). The delayed response was not due to autocrine cytokine secretion with subsequent autostimulation of the secreting cells or to UV-induced growth factor receptor activation. Cells unable to repair UVC-induced DNA damage responded to lower doses of UVC with an even greater accumulation of c-fos and Kin17 mRNAs than repair-proficient wild-type cells, suggesting that a process in which a repair protein is involved regulates mRNA stability. Although resembling the induction of p53, a DNA damage-dependent increase in p53 was not a necessary intermediate in the stabilization reaction, since cells derived from p53 knockout mice showed the same pattern of c-fos and Kin17 mRNA accumulation as wild-type cells. The data indicate that the signal flow induced by UV radiation addresses not only protein stability (p53) and transcription but also RNA stability, a hitherto-unrecognized level of UV-induced regulation.
Figures

References
-
- Adler V, Fuchs S Y, Kim J, Kraft A, King M P, Pelling J, Ronai Z. Jun-NH2-terminal kinase activation mediated by UV-induced DNA lesions in melanoma and fibroblast cells. Cell Growth Differ. 1995;6:1437–1446. - PubMed
-
- Angulo J F, Moreau P L, Maunoury R, Laporte J, Hill A M, Bertolotti R, Devoret R. KIN, a mammalian nuclear protein immunologically related to E. coli RecA protein. Mutat Res. 1989;217:123–134. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous