

Complex transcriptional regulatory mechanisms control expression of the E2F3 locus
- PMID: 10779353
- PMCID: PMC85656
- DOI: 10.1128/MCB.20.10.3633-3639.2000
Complex transcriptional regulatory mechanisms control expression of the E2F3 locus
Abstract
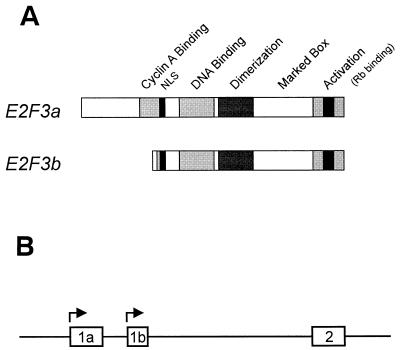
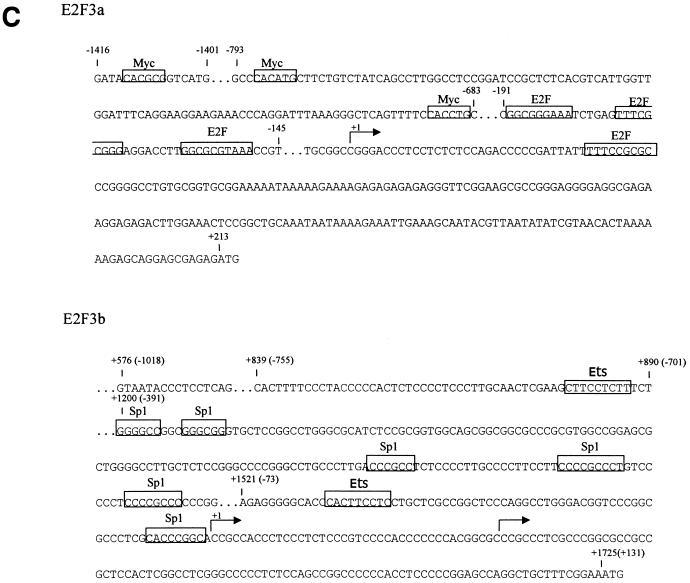
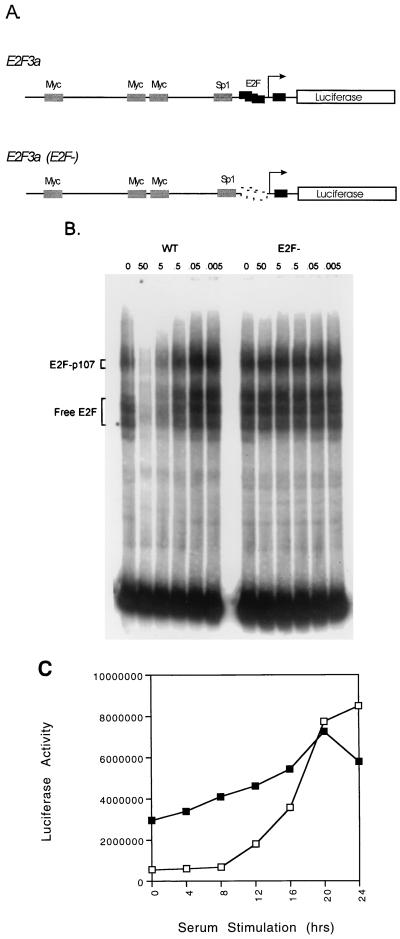
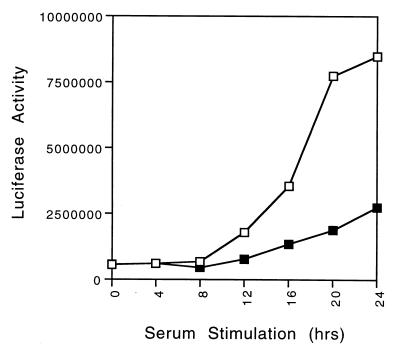
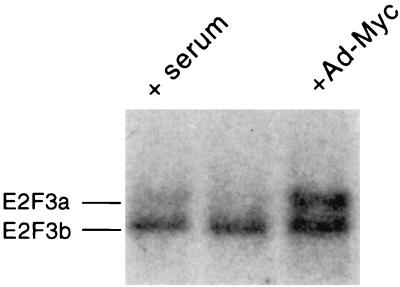
E2F transcription activity has been shown to play a critical role in cell growth control, regulating the expression of a variety of genes that encode proteins important for the initiation of DNA replication and cell cycle regulation. We have shown that the E2F3 locus encodes two protein products: the E2F3a product, which is tightly regulated by cell growth, and the E2F3b product, which is constitutively expressed throughout the cell cycle. To further explore the mechanism controlling the expression of the two E2F3 gene products, we analyzed the genomic sequences flanking the 5' region of E2F3a and E2F3b. We find that a series of E2F binding sites confer negative control on the E2F3a promoter in quiescent cells, similar to the control of the E2F1 and E2F2 promoters. In addition, a group of E-box elements, which are Myc binding sites, confer responsiveness to Myc and are necessary for full activation of the E2F3a promoter in response to growth stimulation. Based on these results and past experiments, it appears that the E2F1, E2F2, and E2F3a genes are similarly regulated by growth stimulation, involving a combination of E2F-dependent negative control and Myc-mediated positive control. In contrast, the constitutive expression of the E2F3b gene more closely reflects the control of expression of the E2F4 and E2F5 genes.
Figures


References
-
- Beijersbergen R L, Kerkhoven R M, Zhu L, Carlee L, Voorhoeve P M, Bernards R. E2F-4, a new member of the E2F gene family, has oncogenic activity and associates with p107 in vivo. Genes Dev. 1994;8:2680–2690. - PubMed
-
- Dyson N. The regulation of E2f by pRB-family proteins. Genes Dev. 1998;12:2245–2262. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous