

ADP-ribosylation factor 6 regulates actin cytoskeleton remodeling in coordination with Rac1 and RhoA
- PMID: 10779358
- PMCID: PMC85663
- DOI: 10.1128/MCB.20.10.3685-3694.2000
ADP-ribosylation factor 6 regulates actin cytoskeleton remodeling in coordination with Rac1 and RhoA
Abstract
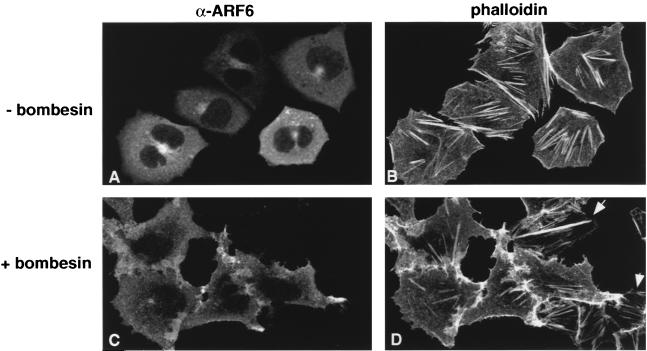
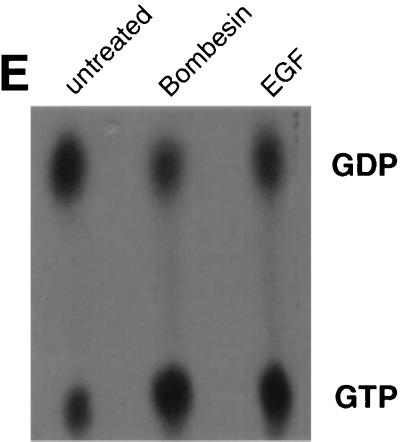
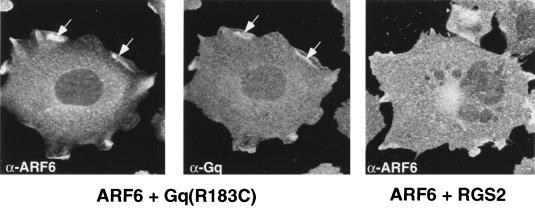
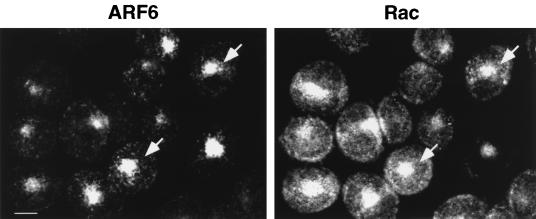
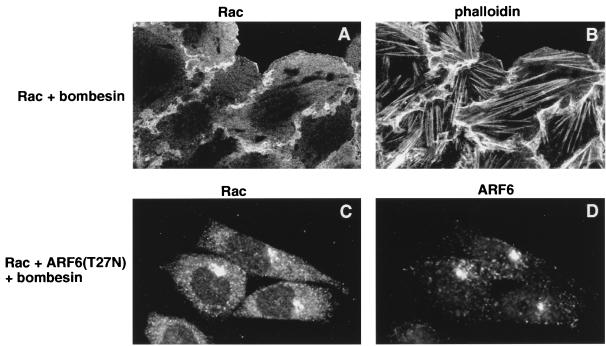
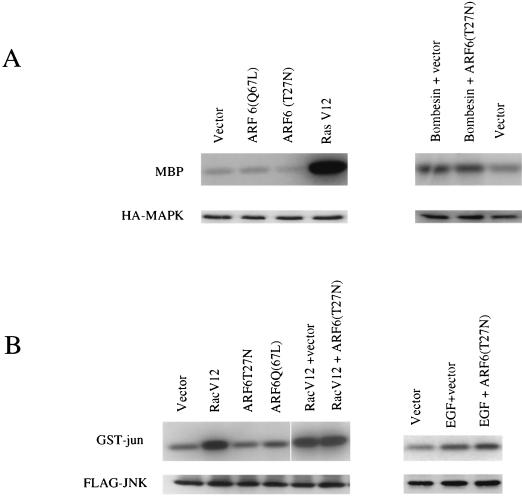
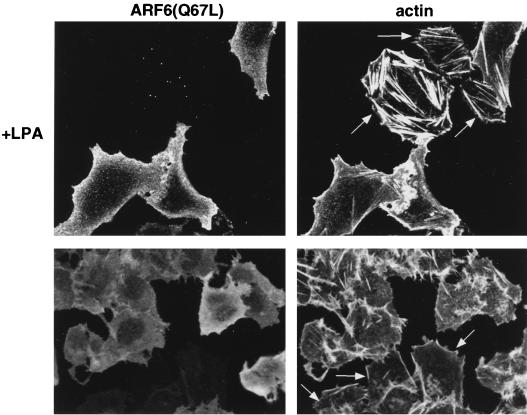
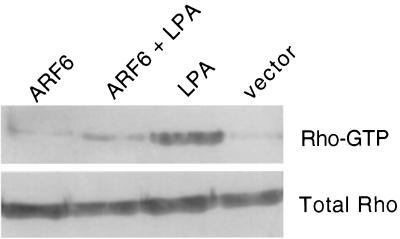
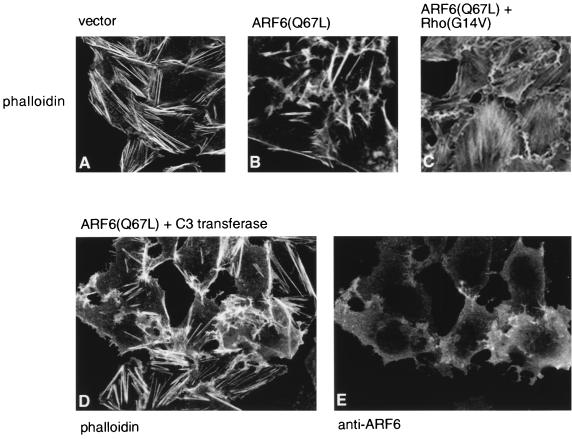
In this study, we have documented an essential role for ADP-ribosylation factor 6 (ARF6) in cell surface remodeling in response to physiological stimulus and in the down regulation of stress fiber formation. We demonstrate that the G-protein-coupled receptor agonist bombesin triggers the redistribution of ARF6- and Rac1-containing endosomal vesicles to the cell surface. This membrane redistribution was accompanied by cortical actin rearrangements and was inhibited by dominant negative ARF6, implying that bombesin is a physiological trigger of ARF6 activation. Furthermore, these studies provide a new model for bombesin-induced Rac1 activation that involves ARF6-regulated endosomal recycling. The bombesin-elicited translocation of vesicular ARF6 was mimicked by activated Galphaq and was partially inhibited by expression of RGS2, which down regulates Gq function. This suggests that Gq functions as an upstream regulator of ARF6 activation. The ARF6-induced peripheral cytoskeletal rearrangements were accompanied by a depletion of stress fibers. Moreover, cells expressing activated ARF6 resisted the formation of stress fibers induced by lysophosphatidic acid. We show that the ARF6-dependent inhibition of stress fiber formation was due to an inhibition of RhoA activation and was overcome by expression of a constitutively active RhoA mutant. The latter observations demonstrate that activation of ARF6 down regulates Rho signaling. Our findings underscore the potential roles of ARF6, Rac1, and RhoA in the coordinated regulation of cytoskeletal remodeling.
Figures
References
-
- Aprikian A G, Tremblay L, Han K, Chevalier S. Bombesin stimulates the motility of human prostate-carcinoma cells through tyrosine phosphorylation of focal adhesion kinase and of integrin-associated proteins. Int J Cancer. 1997;72:498–504. - PubMed
-
- Bretscher M S. Getting membrane flow and the cytoskeleton to cooperate in moving cells. Cell. 1996;87:601–606. - PubMed
-
- Bretscher M S, Aguado-Velasco C. EGF induces recycling membrane to form ruffles. Curr Biol. 1998;8:721–724. - PubMed
-
- Brown H A, Gutowski S, Moomaw C R, Slaughter C, Sternweiss P C. ADP-ribosylation factor, a small GTP-dependent regulatory protein stimulates phospholipase D activity. Cell. 1993;75:1137–1144. - PubMed
-
- Charlesworth A, Rozengurt E. Bombesin and neuromedin B stimulate the activation of p42(mapk) and p74(raf-1) via a protein kinase C-independent pathway in Rat-1 cells. Oncogene. 1997;14:2323–2329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials