

Review
doi: 10.1073/pnas.97.9.4434.
Evolution of the bilaterian body plan: what have we learned from annelids?
Collaborators,
Affiliations
- PMID: 10781038
- PMCID: PMC34316
- DOI: 10.1073/pnas.97.9.4434
Item in Clipboard
Review
Evolution of the bilaterian body plan: what have we learned from annelids?
Proc Natl Acad Sci U S A.
.
Abstract
Annelids, unlike their vertebrate or fruit fly cousins, are a bilaterian taxon often overlooked when addressing the question of body plan evolution. However, recent data suggest that annelids offer unique insights on the early evolution of spiral cleavage, anteroposterior axis formation, body axis segmentation, and head versus trunk distinction.
Figures
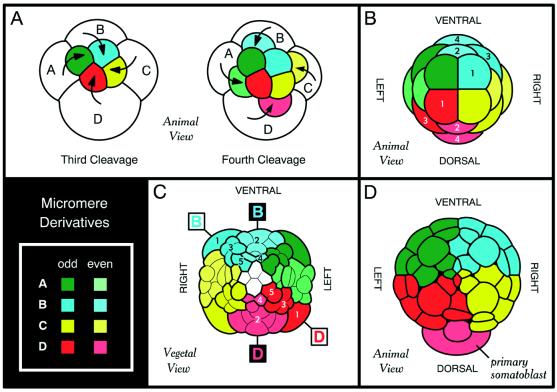
Annelids and a number of other lophotrochozoans manifest a conserved
pattern of early development known as spiral cleavage.
(A) The first two cleavage planes fall at right angles
parallel to the animal–vegetal axis and divide the zygote into the A,
B, C, and D quadrants. In some but not all spiralians, the D blastomere
is larger than the rest. Beginning with the third round of cleavage,
the A, B, C, and D blastomeres cleave off (arrows) quartets of smaller
cells called micromeres at the animal pole. Micromeres are colored
according to quadrant of origin, with color intensity differing for
odd- and even-numbered quartets. In spiral cleavage, each quartet of
micromeres is rotated with respect to the parent blastomere, and the
chirality of rotation alternates for odd- and even-numbered quartets.
(B) Embryonic fate map of the nemertean
Cerebratulus (adapted from ref. 4). Clones derived from
the four B quadrant micromeres (cyan) and D quadrant micromeres (red)
are numbered. Note that odd- and even-numbered quartets have distinct
symmetry properties, with the odd-numbered micromeres being rotated
45° clockwise as viewed from the animal pole. Thus, in the first and
third quartets, the A and D quadrants are on the left, bilaterally
symmetrical to the B and C quadrants on the right. Only the second and
fourth quartets have the “traditional” spiralian fate map (1),
with D being dorsal and B ventral. (C and
D) Although ignored for many years, the alternating
symmetry of the spiralian fate map is readily apparent in the tracings
of early annelid embryologists. C is an adaptation of R.
Woltereck's (30) tracing of the polychaete annelid
Polygordius nearing the end of gastrulation. Thick
outlines demarcate clones derived from single micromeres. Clones
derived from the five B quadrant micromeres (cyan) and D quadrant
micromeres (red) are numbered, and it can be seen that the plane
bisecting the B and D quadrants is rotated by 45° for odd- and
even-numbered quartets. Part D is an adaptation of E. B. Wilson's
(2) tracing of the polychaete annelid Nereis at a
similar stage but seen from the animal pole. The animal hemisphere is
composed of the four primary micromere clones (same color scheme as
other figures), with the D lineage contributing to the left-dorsal
quadrant. Also note that the second quartet micromere from the D
lineage (primary somatoblast) straddles the dorsal midline. The primary
and secondary somatoblasts (second and fourth quartet micromeres from
the D lineage) are the main source of ectoderm and mesoderm in the
adult spiralian body plan, and in Nereis, these cells
are larger than the other micromeres. The symmetry properties of these
two even-numbered micromeres became an all-encompassing tenet of
spiralian embryology (i.e., D is dorsal) for most of the 20th century,
and it is only with the advent of modern cell-labeling techniques that
the true complexity of the spiralian fate map has been rediscovered
(–8).
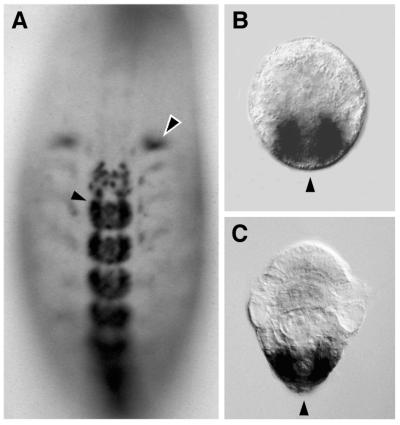
Expression of Hox genes in developing annelids shown by in
situ hybridization. (A) In embryos of the leech
Helobdella triserialis, expression of the Hox genes
begins during organogenesis, long after the formation of segments and
the specification of segment identity (10, 14, 15). Embryo is stained
for expression of Lox2 (Hox paralogue group 7/8) and
shown in ventral view with anterior to the top of the page.
Lox2 RNA is detected only in the posterior two-thirds of
the body plan, including intense staining in the ganglia of the central
nervous system (solid arrow) and reproductive structures (hollow arrow)
and faint staining in the segmental mesoderm. (B and
C) The onset of Hox gene expression in larvae of the
polychaete annelid Chaetopterus varieopedatus is
coincident with the formation of segments and first appears in a
posterior growth zone from which the differentiating segments emerge
(11). Larvae are shown in dorsal view with anterior to the top of the
page and an arrowhead marking the posterior pole. Gene expression is
restricted to the posterior growth zone, on either side of the pole.
B shows expression of gene CHv-Hox2 (Hox
paralogue group 2), and C shows expression of gene
CHv-Hox3 (Hox paralogue group 3). Images in
B and C are courtesy of Steve Irvine and
Mark Martindale (Univ. of Hawaii, Honolulu, HI).
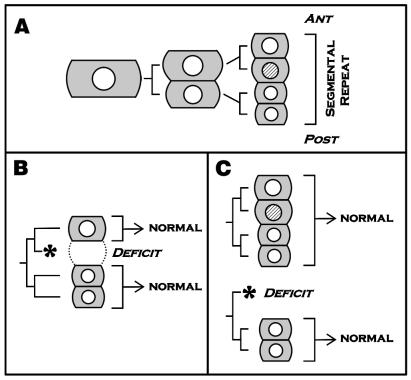
Expression of the en gene does not seem to be required
for the establishment of normal segment polarity in the leech
Helobdella (ref. and E.C.S. and M.S., unpublished
results). (A) The primary p blast cell gives rise to one
segmental repeat of the leech's dorsolateral ectoderm. The en protein
(shaded nucleus) is expressed in only one of the four granddaughters of
the primary blast cell (21). ANT, anterior; POST, posterior.
(B) Laser ablation of the en-expressing
cell has no detectable effect on the specification of more anterior or
posterior parts of that same blast cell clone. (C) Laser
ablation of the anterior daughter of the primary P blast cell prevents
the formation of the en-expressing granddaughter. This
manipulation has no detectable effect on the specification of the
posterior half of that same blast cell clone nor on the segment
polarity of the next anterior blast cell clone. These results suggest
that en-initiated cell interactions are not required for
the proper specification of segment polarity in the leech.
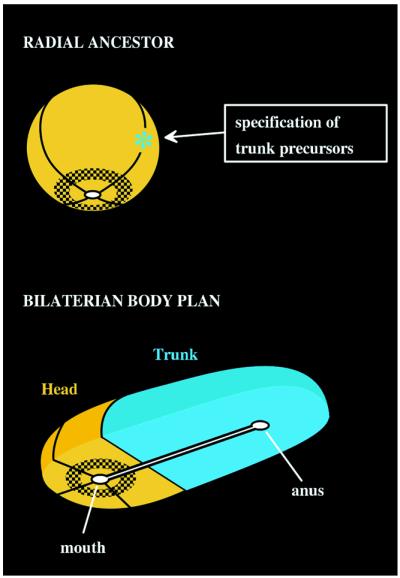
Radial head model for the origin of the bilaterian body plan.
(A) This model assumes a prebilaterian ancestor (orange)
with a radially organized body plan and a single gut opening, shown
here at the bottom. For convenience, the ancestral body plan has been
divided into quarters, and a hypothetical gene expression domain is
shown by shading. Transition to the modern bilaterian body plan began
with the asymmetric specification of a specialized group of
“trunk” precursor cells (cyan) at only one meridian around the
circumference of the ancestral body plan. (B) Allometric
expansion of the trunk domain produces a body plan typical of most
Bilateria. The trunk elongates away from the head domain, carrying with
it the anal end of a now bipolar gut. But the head domain retains
features of its ancestral radial organization, as noted by a gene
expression pattern (shaded) concentric around the mouth. This model
proposes that bilaterian “head genes” have been relegated to the
head domain, because they were not coopted into trunk patterning, and
suggests that the AP axis may be an innovation of the Bilateria rather
than a modification of a preexisting axis. [Reproduced with permission
from ref. (Copyright 1998, Academic Press)].
References
-
- Brusca R C, Brusca G J. Invertebrates. Sunderland, MA: Sinauer; 1990.
-
- Wilson E B. J Morphol. 1892;6:361–480.
-
- Adoutte A, Balavoine G, Lartillot N, de Rosa R. Trends Genet. 1999;15:104–108. - PubMed
-
- Henry J J, Martindale M Q. Dev Biol. 1998;201:253–269. - PubMed
-
- Boyer B C, Henry J J, Martindale M Q. Dev Biol. 1998;204:111–123. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
