

Specific and heritable genetic interference by double-stranded RNA in Arabidopsis thaliana
- PMID: 10781109
- PMCID: PMC18344
- DOI: 10.1073/pnas.060034297
Specific and heritable genetic interference by double-stranded RNA in Arabidopsis thaliana
Abstract
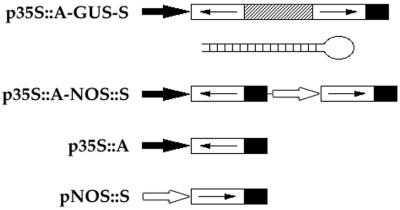
We investigated the potential of double-stranded RNA interference (RNAi) with gene activity in Arabidopsis thaliana. To construct transformation vectors that produce RNAs capable of duplex formation, gene-specific sequences in the sense and antisense orientations were linked and placed under the control of a strong viral promoter. When introduced into the genome of A. thaliana by Agrobacterium-mediated transformation, double-stranded RNA-expressing constructs corresponding to four genes, AGAMOUS (AG), CLAVATA3, APETALA1, and PERIANTHIA, caused specific and heritable genetic interference. The severity of phenotypes varied between transgenic lines. In situ hybridization revealed a correlation between a declining AG mRNA accumulation and increasingly severe phenotypes in AG (RNAi) mutants, suggesting that endogenous mRNA is the target of double-stranded RNA-mediated genetic interference. The ability to generate stably heritable RNAi and the resultant specific phenotypes allows us to selectively reduce gene function in A. thaliana.
Figures





References
-
- Coles J P, Phillips A L, Croker S J, Garcia-Lepe R, Lewis M J, Hedden P. Plant J. 1999;17:547–556. - PubMed
-
- Haldrup A, Naver H, Scheller H V. Plant Cell. 1999;17:689–698. - PubMed
-
- Nanjo T, Kobayashi M, Yoshiba Y, Sanada Y, Wada K, Tsukaya H, Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K. Plant J. 1999;18:185–193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
