

The leucine-rich repeat protein SUR-8 enhances MAP kinase activation and forms a complex with Ras and Raf
Affiliations
- PMID: 10783161
- PMCID: PMC316541
Item in Clipboard
The leucine-rich repeat protein SUR-8 enhances MAP kinase activation and forms a complex with Ras and Raf
Genes Dev.
.
Abstract
Caenorhabditis elegans sur-8 encodes a positive regulator of Ras signaling. We investigated the mechanism by which the human Sur-8 homolog can positively regulate Ras-MAP kinase signaling in mammalian cells. Sur-8 expression enhances Ras- or EGF-induced Raf and ERK activation but has no effect on ERK activation induced by active Raf or MEK. Furthermore, Sur-8 expression does not increase AKT or JNK activation. Sur-8 interacts with Ras and Raf and is able to form a ternary complex with the two proteins. Thus, Sur-8 may function as a scaffold that enhances Ras-MAP kinase signal transduction by facilitating the interaction between Ras and Raf.
Figures
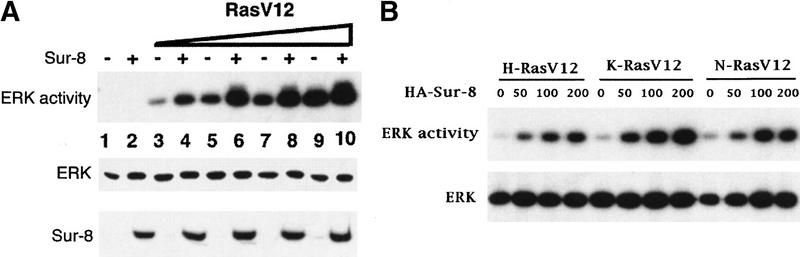
Sur-8 enhances ERK activation. (A) Myc-tagged ERK1 (200 ng) was cotransfected with increasing amounts of RasV12 in the presence or absence of Sur-8 (500 ng) as indicated. The amounts of RasV12 (all Ras plasmids used are K-Ras except as described) used for transfection are 0 (lanes 1,2), 5 ng (lanes 3,4), 10 ng (lanes 5,6), 20 ng (lanes 7,8), and 50 ng (lanes 9,10). Myc–ERK was immunoprecipitated and kinase activity was determined using GST–Elk-1 as a substrate (top). Myc–ERK in the immunoprecipitate was determined by anti-ERK Western blot (middle). Expression of HA–Sur-8 in cell lysates was detected by anti-HA Western blot (bottom). (B) Dose-dependent effect of Sur-8 on ERK activity. Cells were cotransfected with 1 ng of H-RasV12, K-RasV12, or N-RasV12, and 200 ng of Myc–ERK. The amount of HA–Sur-8 (ng DNA) cotransfected is indicated. (C) Sur-8 enhances EGF-stimulated ERK activity. The 293 cells were cotransfected with 200 ng of Myc–ERK and 500 ng of HA–Sur-8. After cells were starved in serum-free medium overnight, the cells were stimulated for 5 min by indicated concentrations of EGF. Myc–ERK was immunoprecipitated and activity was determined. (D) Mutations at Cys-260 and Glu457 of Sur-8 compromised its functions in ERK activation. Sur-8 or mutants (50 and 500 ng) were cotransfected with Myc–ERK and RasV12. Immunoprecipitated ERK activity and expression levels are shown at top and middle, respectively. HA–Sur-8 expression was detected as above (bottom).
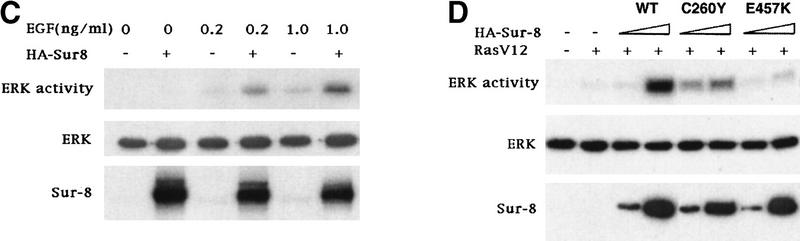
Sur-8 enhances ERK activation. (A) Myc-tagged ERK1 (200 ng) was cotransfected with increasing amounts of RasV12 in the presence or absence of Sur-8 (500 ng) as indicated. The amounts of RasV12 (all Ras plasmids used are K-Ras except as described) used for transfection are 0 (lanes 1,2), 5 ng (lanes 3,4), 10 ng (lanes 5,6), 20 ng (lanes 7,8), and 50 ng (lanes 9,10). Myc–ERK was immunoprecipitated and kinase activity was determined using GST–Elk-1 as a substrate (top). Myc–ERK in the immunoprecipitate was determined by anti-ERK Western blot (middle). Expression of HA–Sur-8 in cell lysates was detected by anti-HA Western blot (bottom). (B) Dose-dependent effect of Sur-8 on ERK activity. Cells were cotransfected with 1 ng of H-RasV12, K-RasV12, or N-RasV12, and 200 ng of Myc–ERK. The amount of HA–Sur-8 (ng DNA) cotransfected is indicated. (C) Sur-8 enhances EGF-stimulated ERK activity. The 293 cells were cotransfected with 200 ng of Myc–ERK and 500 ng of HA–Sur-8. After cells were starved in serum-free medium overnight, the cells were stimulated for 5 min by indicated concentrations of EGF. Myc–ERK was immunoprecipitated and activity was determined. (D) Mutations at Cys-260 and Glu457 of Sur-8 compromised its functions in ERK activation. Sur-8 or mutants (50 and 500 ng) were cotransfected with Myc–ERK and RasV12. Immunoprecipitated ERK activity and expression levels are shown at top and middle, respectively. HA–Sur-8 expression was detected as above (bottom).
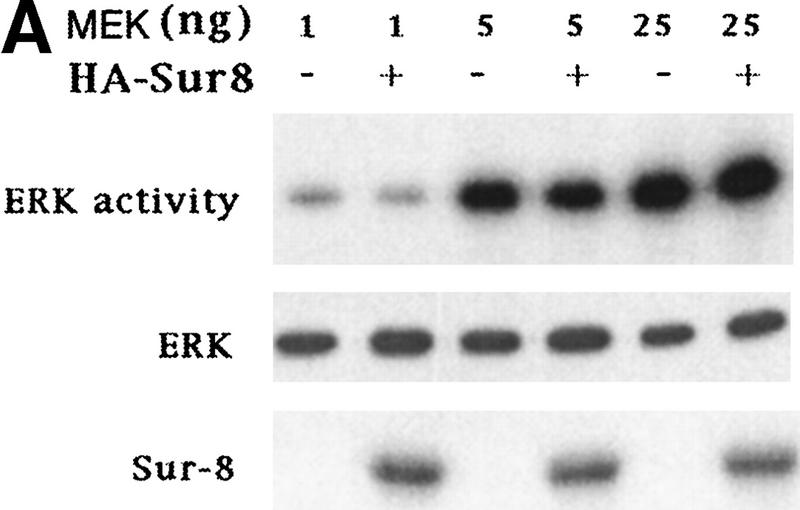
Sur-8 specifically enhances the ERK pathway and functions upstream of Raf. (A) Sur-8 has no effect on MEK-induced ERK activation. The 293 cells were cotransfected with 500 ng of HA–Sur-8, 200 ng of Myc–ERK and varied amounts of a constitutively active MEK mutant as indicated. ERK activity was determined as in Fig. 1A. (B) Sur-8 has no effect on Raf-induced ERK activation. Myc–ERK (200 ng) was cotransfected with V-Raf (10 ng, left) or varied amounts of active B-Raf mutant (right). Cotransfection of Sur-8 did not further increase ERK activity. (C) Sur-8 enhances Raf activity in response to RasV12 or EGF stimulation. 293 cells were cotransfected with 400 ng of Flag–Raf, 20 ng of HA–RasV12, and varied amounts of HA–Sur-8 as indicated. Cells were stimulated with EGF for 3 min where indicated. Flag–Raf was immunoprecipitated with anti-Flag and activity was determined by the coupled kinase assay. Expression of Sur-8 and Ras in lysates and the levels of Raf in the immunoprecipitates were confirmed by Western blot. (D) Sur-8 has no effect on JNK activation. HA–JNK (300 ng) was transfected into cells. Cotransfection with RasV12 or treatment with anisomycin was used to stimulate JNK activity. HA–JNK was immunoprecipitated and assayed for kinase activity using GST–Jun as a substrate. (E) RasV12-induced AKT activation is not enhanced by Sur-8. Wild-type GST–AKT (100 ng) was cotransfected into cells with HA–RasV12 and varied amounts of Sur-8 as indicated. GST–AKT were purified and kinase activity was determined using histone 2B as a substrate. (Lane 7) Negative control of the kinase dead mutant (KD) of GST–AKT.
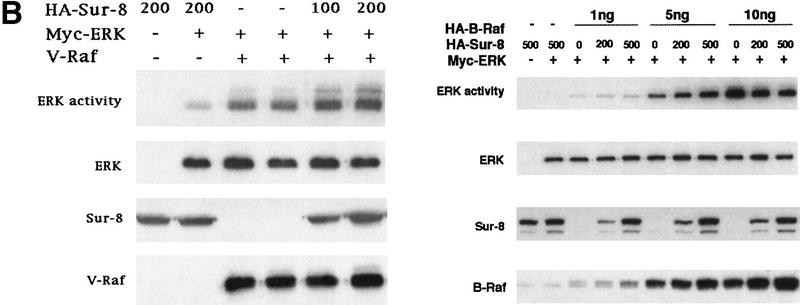
Sur-8 specifically enhances the ERK pathway and functions upstream of Raf. (A) Sur-8 has no effect on MEK-induced ERK activation. The 293 cells were cotransfected with 500 ng of HA–Sur-8, 200 ng of Myc–ERK and varied amounts of a constitutively active MEK mutant as indicated. ERK activity was determined as in Fig. 1A. (B) Sur-8 has no effect on Raf-induced ERK activation. Myc–ERK (200 ng) was cotransfected with V-Raf (10 ng, left) or varied amounts of active B-Raf mutant (right). Cotransfection of Sur-8 did not further increase ERK activity. (C) Sur-8 enhances Raf activity in response to RasV12 or EGF stimulation. 293 cells were cotransfected with 400 ng of Flag–Raf, 20 ng of HA–RasV12, and varied amounts of HA–Sur-8 as indicated. Cells were stimulated with EGF for 3 min where indicated. Flag–Raf was immunoprecipitated with anti-Flag and activity was determined by the coupled kinase assay. Expression of Sur-8 and Ras in lysates and the levels of Raf in the immunoprecipitates were confirmed by Western blot. (D) Sur-8 has no effect on JNK activation. HA–JNK (300 ng) was transfected into cells. Cotransfection with RasV12 or treatment with anisomycin was used to stimulate JNK activity. HA–JNK was immunoprecipitated and assayed for kinase activity using GST–Jun as a substrate. (E) RasV12-induced AKT activation is not enhanced by Sur-8. Wild-type GST–AKT (100 ng) was cotransfected into cells with HA–RasV12 and varied amounts of Sur-8 as indicated. GST–AKT were purified and kinase activity was determined using histone 2B as a substrate. (Lane 7) Negative control of the kinase dead mutant (KD) of GST–AKT.
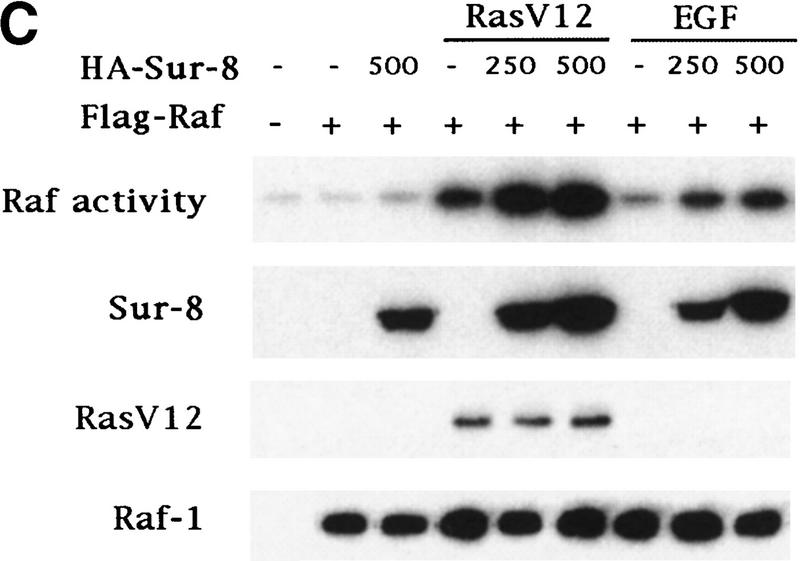
Sur-8 specifically enhances the ERK pathway and functions upstream of Raf. (A) Sur-8 has no effect on MEK-induced ERK activation. The 293 cells were cotransfected with 500 ng of HA–Sur-8, 200 ng of Myc–ERK and varied amounts of a constitutively active MEK mutant as indicated. ERK activity was determined as in Fig. 1A. (B) Sur-8 has no effect on Raf-induced ERK activation. Myc–ERK (200 ng) was cotransfected with V-Raf (10 ng, left) or varied amounts of active B-Raf mutant (right). Cotransfection of Sur-8 did not further increase ERK activity. (C) Sur-8 enhances Raf activity in response to RasV12 or EGF stimulation. 293 cells were cotransfected with 400 ng of Flag–Raf, 20 ng of HA–RasV12, and varied amounts of HA–Sur-8 as indicated. Cells were stimulated with EGF for 3 min where indicated. Flag–Raf was immunoprecipitated with anti-Flag and activity was determined by the coupled kinase assay. Expression of Sur-8 and Ras in lysates and the levels of Raf in the immunoprecipitates were confirmed by Western blot. (D) Sur-8 has no effect on JNK activation. HA–JNK (300 ng) was transfected into cells. Cotransfection with RasV12 or treatment with anisomycin was used to stimulate JNK activity. HA–JNK was immunoprecipitated and assayed for kinase activity using GST–Jun as a substrate. (E) RasV12-induced AKT activation is not enhanced by Sur-8. Wild-type GST–AKT (100 ng) was cotransfected into cells with HA–RasV12 and varied amounts of Sur-8 as indicated. GST–AKT were purified and kinase activity was determined using histone 2B as a substrate. (Lane 7) Negative control of the kinase dead mutant (KD) of GST–AKT.
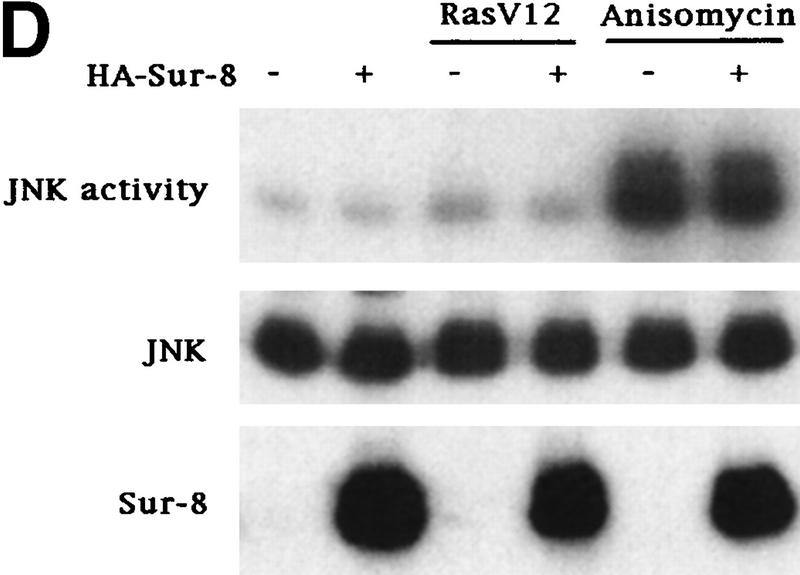
Sur-8 specifically enhances the ERK pathway and functions upstream of Raf. (A) Sur-8 has no effect on MEK-induced ERK activation. The 293 cells were cotransfected with 500 ng of HA–Sur-8, 200 ng of Myc–ERK and varied amounts of a constitutively active MEK mutant as indicated. ERK activity was determined as in Fig. 1A. (B) Sur-8 has no effect on Raf-induced ERK activation. Myc–ERK (200 ng) was cotransfected with V-Raf (10 ng, left) or varied amounts of active B-Raf mutant (right). Cotransfection of Sur-8 did not further increase ERK activity. (C) Sur-8 enhances Raf activity in response to RasV12 or EGF stimulation. 293 cells were cotransfected with 400 ng of Flag–Raf, 20 ng of HA–RasV12, and varied amounts of HA–Sur-8 as indicated. Cells were stimulated with EGF for 3 min where indicated. Flag–Raf was immunoprecipitated with anti-Flag and activity was determined by the coupled kinase assay. Expression of Sur-8 and Ras in lysates and the levels of Raf in the immunoprecipitates were confirmed by Western blot. (D) Sur-8 has no effect on JNK activation. HA–JNK (300 ng) was transfected into cells. Cotransfection with RasV12 or treatment with anisomycin was used to stimulate JNK activity. HA–JNK was immunoprecipitated and assayed for kinase activity using GST–Jun as a substrate. (E) RasV12-induced AKT activation is not enhanced by Sur-8. Wild-type GST–AKT (100 ng) was cotransfected into cells with HA–RasV12 and varied amounts of Sur-8 as indicated. GST–AKT were purified and kinase activity was determined using histone 2B as a substrate. (Lane 7) Negative control of the kinase dead mutant (KD) of GST–AKT.
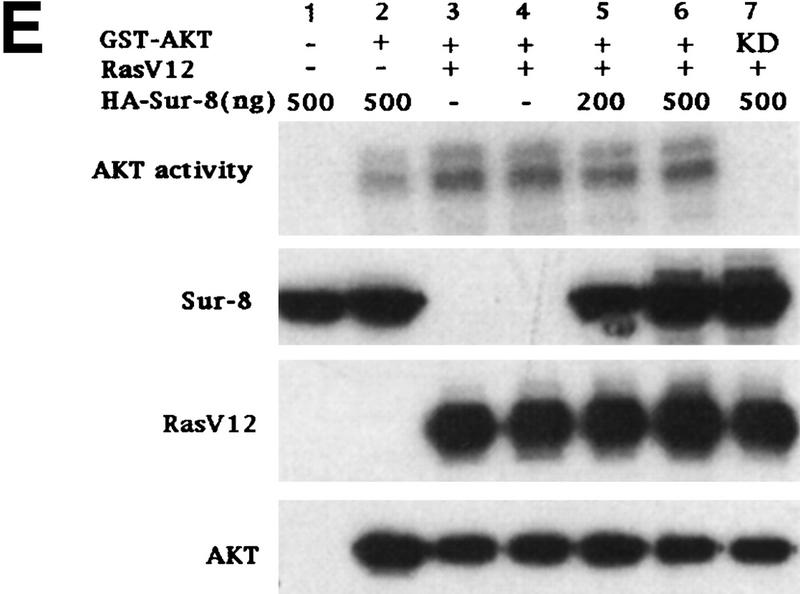
Sur-8 specifically enhances the ERK pathway and functions upstream of Raf. (A) Sur-8 has no effect on MEK-induced ERK activation. The 293 cells were cotransfected with 500 ng of HA–Sur-8, 200 ng of Myc–ERK and varied amounts of a constitutively active MEK mutant as indicated. ERK activity was determined as in Fig. 1A. (B) Sur-8 has no effect on Raf-induced ERK activation. Myc–ERK (200 ng) was cotransfected with V-Raf (10 ng, left) or varied amounts of active B-Raf mutant (right). Cotransfection of Sur-8 did not further increase ERK activity. (C) Sur-8 enhances Raf activity in response to RasV12 or EGF stimulation. 293 cells were cotransfected with 400 ng of Flag–Raf, 20 ng of HA–RasV12, and varied amounts of HA–Sur-8 as indicated. Cells were stimulated with EGF for 3 min where indicated. Flag–Raf was immunoprecipitated with anti-Flag and activity was determined by the coupled kinase assay. Expression of Sur-8 and Ras in lysates and the levels of Raf in the immunoprecipitates were confirmed by Western blot. (D) Sur-8 has no effect on JNK activation. HA–JNK (300 ng) was transfected into cells. Cotransfection with RasV12 or treatment with anisomycin was used to stimulate JNK activity. HA–JNK was immunoprecipitated and assayed for kinase activity using GST–Jun as a substrate. (E) RasV12-induced AKT activation is not enhanced by Sur-8. Wild-type GST–AKT (100 ng) was cotransfected into cells with HA–RasV12 and varied amounts of Sur-8 as indicated. GST–AKT were purified and kinase activity was determined using histone 2B as a substrate. (Lane 7) Negative control of the kinase dead mutant (KD) of GST–AKT.

Sur-8 interacts with Ras and Raf. (A) Interactions between Sur-8 and Ras mutants in the yeast two-hybrid assays. Growth on SC-HLW medium indicates a positive interaction. RasY32F and RasE37G are in the RasV12 mutant background. (B) Coimmunoprecipitation between Sur-8 and RasV12. A total of 500 ng of HA–Sur-8 was cotransfected into 293 cells with 200 ng of RasV12 as indicated. Thirty-six hours after transfection, cells lysates were immunoprecipitated with anti-Ras antibody (left) or anti-HA (right) as indicated. The immunoprecipitates were blotted with anti-HA (left) or anti-Ras (right). (Lane L) Cell lysate controls. (IP, IB) Immunoprecipitation and immunoblot, respectively. (IgG) The immunoglobulin band. (C) Interaction between Sur-8 and Raf were determined by coimmunoprecipitation of transfected HA–Sur-8 and Myc–Raf. (Lane L) The cell lysate control. (D) The carboxy-terminal domain of Raf interacts with Sur-8. Flag-tagged Raf full length, ΔN (residues 325–648), ΔC (residues 1–256) were cotransfected with HA–Sur-8. HA–Sur-8 was immunoprecipitated by anti-HA followed by Western blot with anti-Flag antibody. Negative controls without anti-HA antibody were included. Bands corresponding to full length, ΔC, and ΔN of Raf are indicated. (E) Coimmunoprecipitation between endogenous Sur-8 and Raf. The 293 cell lysates were immunoprecipitated with control serum (lane 1), anti-Ras (lane 2), or anti-Sur-8. The immunoprecipitates were blotted with anti-Raf. (Lane 4) Control of cell lysates.
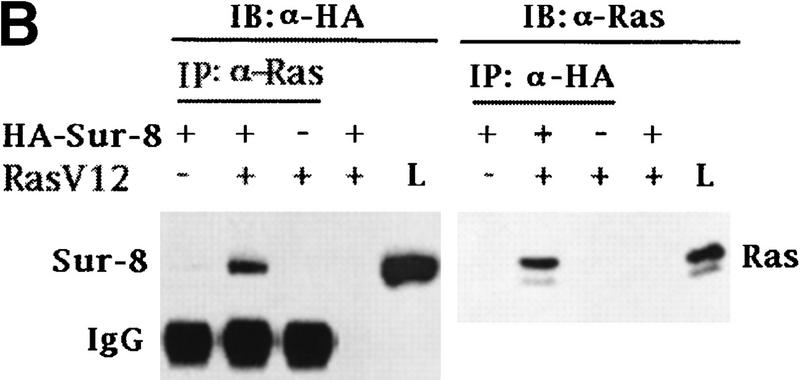
Sur-8 interacts with Ras and Raf. (A) Interactions between Sur-8 and Ras mutants in the yeast two-hybrid assays. Growth on SC-HLW medium indicates a positive interaction. RasY32F and RasE37G are in the RasV12 mutant background. (B) Coimmunoprecipitation between Sur-8 and RasV12. A total of 500 ng of HA–Sur-8 was cotransfected into 293 cells with 200 ng of RasV12 as indicated. Thirty-six hours after transfection, cells lysates were immunoprecipitated with anti-Ras antibody (left) or anti-HA (right) as indicated. The immunoprecipitates were blotted with anti-HA (left) or anti-Ras (right). (Lane L) Cell lysate controls. (IP, IB) Immunoprecipitation and immunoblot, respectively. (IgG) The immunoglobulin band. (C) Interaction between Sur-8 and Raf were determined by coimmunoprecipitation of transfected HA–Sur-8 and Myc–Raf. (Lane L) The cell lysate control. (D) The carboxy-terminal domain of Raf interacts with Sur-8. Flag-tagged Raf full length, ΔN (residues 325–648), ΔC (residues 1–256) were cotransfected with HA–Sur-8. HA–Sur-8 was immunoprecipitated by anti-HA followed by Western blot with anti-Flag antibody. Negative controls without anti-HA antibody were included. Bands corresponding to full length, ΔC, and ΔN of Raf are indicated. (E) Coimmunoprecipitation between endogenous Sur-8 and Raf. The 293 cell lysates were immunoprecipitated with control serum (lane 1), anti-Ras (lane 2), or anti-Sur-8. The immunoprecipitates were blotted with anti-Raf. (Lane 4) Control of cell lysates.
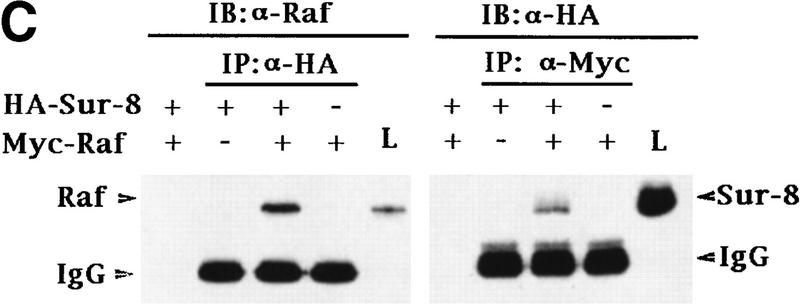
Sur-8 interacts with Ras and Raf. (A) Interactions between Sur-8 and Ras mutants in the yeast two-hybrid assays. Growth on SC-HLW medium indicates a positive interaction. RasY32F and RasE37G are in the RasV12 mutant background. (B) Coimmunoprecipitation between Sur-8 and RasV12. A total of 500 ng of HA–Sur-8 was cotransfected into 293 cells with 200 ng of RasV12 as indicated. Thirty-six hours after transfection, cells lysates were immunoprecipitated with anti-Ras antibody (left) or anti-HA (right) as indicated. The immunoprecipitates were blotted with anti-HA (left) or anti-Ras (right). (Lane L) Cell lysate controls. (IP, IB) Immunoprecipitation and immunoblot, respectively. (IgG) The immunoglobulin band. (C) Interaction between Sur-8 and Raf were determined by coimmunoprecipitation of transfected HA–Sur-8 and Myc–Raf. (Lane L) The cell lysate control. (D) The carboxy-terminal domain of Raf interacts with Sur-8. Flag-tagged Raf full length, ΔN (residues 325–648), ΔC (residues 1–256) were cotransfected with HA–Sur-8. HA–Sur-8 was immunoprecipitated by anti-HA followed by Western blot with anti-Flag antibody. Negative controls without anti-HA antibody were included. Bands corresponding to full length, ΔC, and ΔN of Raf are indicated. (E) Coimmunoprecipitation between endogenous Sur-8 and Raf. The 293 cell lysates were immunoprecipitated with control serum (lane 1), anti-Ras (lane 2), or anti-Sur-8. The immunoprecipitates were blotted with anti-Raf. (Lane 4) Control of cell lysates.
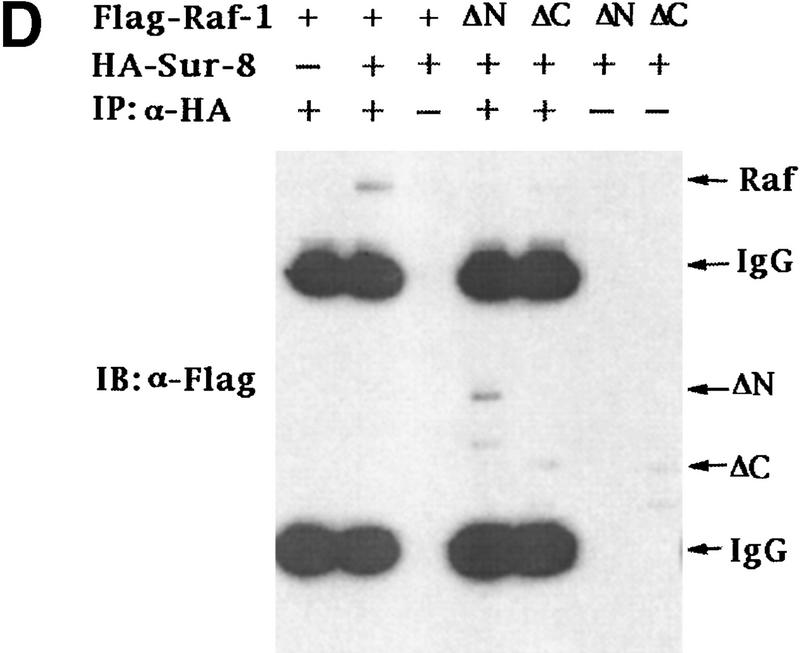
Sur-8 interacts with Ras and Raf. (A) Interactions between Sur-8 and Ras mutants in the yeast two-hybrid assays. Growth on SC-HLW medium indicates a positive interaction. RasY32F and RasE37G are in the RasV12 mutant background. (B) Coimmunoprecipitation between Sur-8 and RasV12. A total of 500 ng of HA–Sur-8 was cotransfected into 293 cells with 200 ng of RasV12 as indicated. Thirty-six hours after transfection, cells lysates were immunoprecipitated with anti-Ras antibody (left) or anti-HA (right) as indicated. The immunoprecipitates were blotted with anti-HA (left) or anti-Ras (right). (Lane L) Cell lysate controls. (IP, IB) Immunoprecipitation and immunoblot, respectively. (IgG) The immunoglobulin band. (C) Interaction between Sur-8 and Raf were determined by coimmunoprecipitation of transfected HA–Sur-8 and Myc–Raf. (Lane L) The cell lysate control. (D) The carboxy-terminal domain of Raf interacts with Sur-8. Flag-tagged Raf full length, ΔN (residues 325–648), ΔC (residues 1–256) were cotransfected with HA–Sur-8. HA–Sur-8 was immunoprecipitated by anti-HA followed by Western blot with anti-Flag antibody. Negative controls without anti-HA antibody were included. Bands corresponding to full length, ΔC, and ΔN of Raf are indicated. (E) Coimmunoprecipitation between endogenous Sur-8 and Raf. The 293 cell lysates were immunoprecipitated with control serum (lane 1), anti-Ras (lane 2), or anti-Sur-8. The immunoprecipitates were blotted with anti-Raf. (Lane 4) Control of cell lysates.
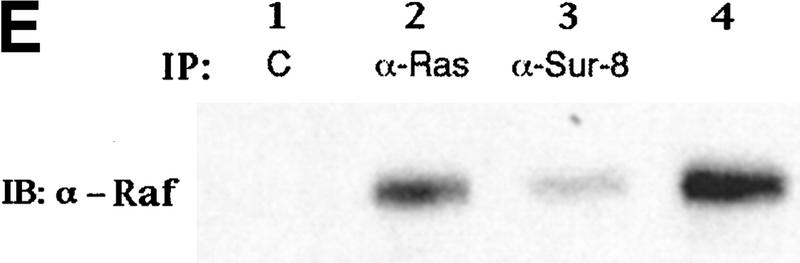
Sur-8 interacts with Ras and Raf. (A) Interactions between Sur-8 and Ras mutants in the yeast two-hybrid assays. Growth on SC-HLW medium indicates a positive interaction. RasY32F and RasE37G are in the RasV12 mutant background. (B) Coimmunoprecipitation between Sur-8 and RasV12. A total of 500 ng of HA–Sur-8 was cotransfected into 293 cells with 200 ng of RasV12 as indicated. Thirty-six hours after transfection, cells lysates were immunoprecipitated with anti-Ras antibody (left) or anti-HA (right) as indicated. The immunoprecipitates were blotted with anti-HA (left) or anti-Ras (right). (Lane L) Cell lysate controls. (IP, IB) Immunoprecipitation and immunoblot, respectively. (IgG) The immunoglobulin band. (C) Interaction between Sur-8 and Raf were determined by coimmunoprecipitation of transfected HA–Sur-8 and Myc–Raf. (Lane L) The cell lysate control. (D) The carboxy-terminal domain of Raf interacts with Sur-8. Flag-tagged Raf full length, ΔN (residues 325–648), ΔC (residues 1–256) were cotransfected with HA–Sur-8. HA–Sur-8 was immunoprecipitated by anti-HA followed by Western blot with anti-Flag antibody. Negative controls without anti-HA antibody were included. Bands corresponding to full length, ΔC, and ΔN of Raf are indicated. (E) Coimmunoprecipitation between endogenous Sur-8 and Raf. The 293 cell lysates were immunoprecipitated with control serum (lane 1), anti-Ras (lane 2), or anti-Sur-8. The immunoprecipitates were blotted with anti-Raf. (Lane 4) Control of cell lysates.
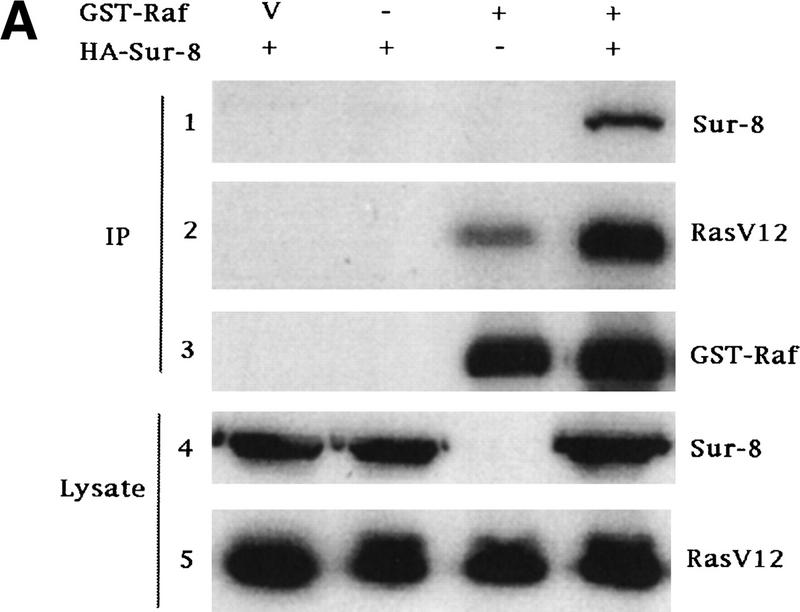
Sur-8 forms a complex with Ras and Raf. (A) Sur-8 enhances the interaction between Ras and Raf. HA–RasV12 was cotransfected with GST–Raf in the presence or absence of HA–Sur-8. GST–Raf was purified by glutathione–agarose resin. The copurified Ras and HA–Sur-8 were detected by anti-HA for HA–Ras (second panel from top) and HA–Sur-8 (top) Western blots. The GST–Raf in the precipitation was detected by an anti-Raf Western blot (middle). Expressions of HA–Sur-8 and HA–RasV12 in cell lysates were shown in bottom two panels. (B) Sur-8C260Y and E457K mutants fail to enhance the interaction between Ras and Raf. Experiments are similar to A. (HA–Sur-8*) Sur-8C260Y in lanes 1–4 and Sur-8E457K in lanes 5–8, respectively. (C) Raf enhances Sur–8-Ras interaction in vitro. Cell lysates expressing HA–Sur-8 or Myc–Raf were mixed with purified GST–RasV12 protein (5 μg). GST–RasV12 and associated proteins were purified by glutathione–agarose and Western blotted with anti-HA to detect HA–Sur-8 (top) and anti-GST to detect GST–RasV12 (bottom). (Lane 6) A control of cell lysate containing HA–Sur-8. (D) Raf bridges the interaction between Ras and Sur-8ΔC. GST–Ras (5 μg) was incubated with cell lysates containing HA–Sur-8ΔC (containing residues 1–366) in the absence (lane 1) or presence (lane 2) of Raf containing cell lysates. HA–Sur-8ΔC (middle) and Raf (top) copurified by GST–Ras were detected by Western blot. GST–Ras coprecipitated HA–Sur-8ΔC only in the presence of Raf. (E) Sur-8, Ras, and Raf form a ternary complex. The 293 cells were transfected with various plasmids as indicated. Cell lysates were subjected to purification by glutathione–agarose beads. The purified proteins were eluted by glutathione and subjected to immunoprecipitation with anti-HA for HA–Ras followed by Western blot with anti-Raf (left). Similarly, proteins eluted from glutathione–agarose beads were immunoprecipitated with anti-Myc for Myc–Raf followed by Western blot with anti-Ras (middle). (Right) Experimental results using His–Sur-8 (lane 8) along with GST–Sur-8 (lane 9). His–Sur-8 complex was first purified by nickel agarose and eluted with imidazole. The eluted samples were immunoprecipitated with anti-Myc and followed by Western blot with anti-Raf.
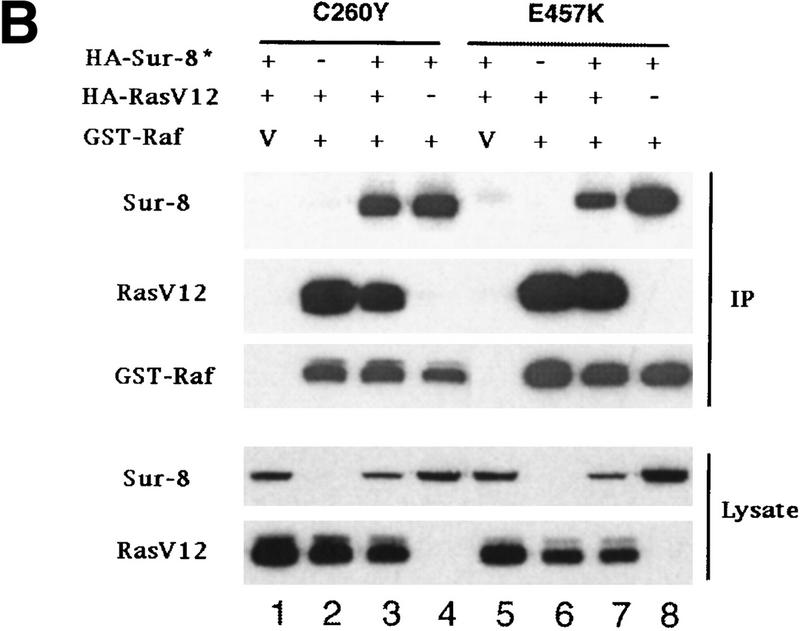
Sur-8 forms a complex with Ras and Raf. (A) Sur-8 enhances the interaction between Ras and Raf. HA–RasV12 was cotransfected with GST–Raf in the presence or absence of HA–Sur-8. GST–Raf was purified by glutathione–agarose resin. The copurified Ras and HA–Sur-8 were detected by anti-HA for HA–Ras (second panel from top) and HA–Sur-8 (top) Western blots. The GST–Raf in the precipitation was detected by an anti-Raf Western blot (middle). Expressions of HA–Sur-8 and HA–RasV12 in cell lysates were shown in bottom two panels. (B) Sur-8C260Y and E457K mutants fail to enhance the interaction between Ras and Raf. Experiments are similar to A. (HA–Sur-8*) Sur-8C260Y in lanes 1–4 and Sur-8E457K in lanes 5–8, respectively. (C) Raf enhances Sur–8-Ras interaction in vitro. Cell lysates expressing HA–Sur-8 or Myc–Raf were mixed with purified GST–RasV12 protein (5 μg). GST–RasV12 and associated proteins were purified by glutathione–agarose and Western blotted with anti-HA to detect HA–Sur-8 (top) and anti-GST to detect GST–RasV12 (bottom). (Lane 6) A control of cell lysate containing HA–Sur-8. (D) Raf bridges the interaction between Ras and Sur-8ΔC. GST–Ras (5 μg) was incubated with cell lysates containing HA–Sur-8ΔC (containing residues 1–366) in the absence (lane 1) or presence (lane 2) of Raf containing cell lysates. HA–Sur-8ΔC (middle) and Raf (top) copurified by GST–Ras were detected by Western blot. GST–Ras coprecipitated HA–Sur-8ΔC only in the presence of Raf. (E) Sur-8, Ras, and Raf form a ternary complex. The 293 cells were transfected with various plasmids as indicated. Cell lysates were subjected to purification by glutathione–agarose beads. The purified proteins were eluted by glutathione and subjected to immunoprecipitation with anti-HA for HA–Ras followed by Western blot with anti-Raf (left). Similarly, proteins eluted from glutathione–agarose beads were immunoprecipitated with anti-Myc for Myc–Raf followed by Western blot with anti-Ras (middle). (Right) Experimental results using His–Sur-8 (lane 8) along with GST–Sur-8 (lane 9). His–Sur-8 complex was first purified by nickel agarose and eluted with imidazole. The eluted samples were immunoprecipitated with anti-Myc and followed by Western blot with anti-Raf.
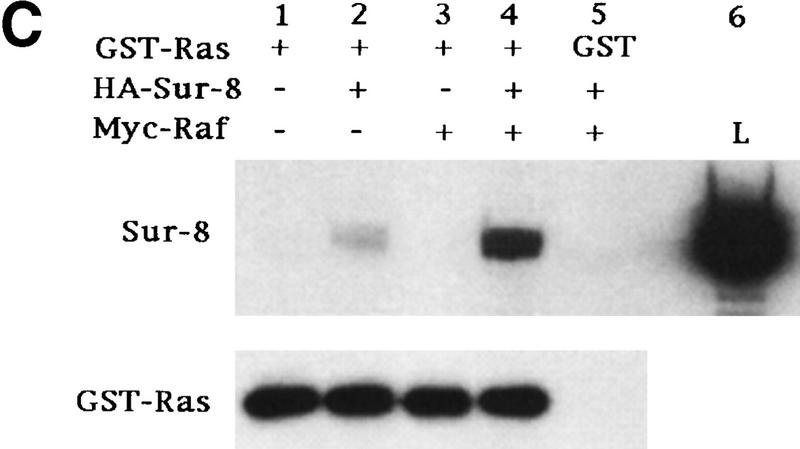
Sur-8 forms a complex with Ras and Raf. (A) Sur-8 enhances the interaction between Ras and Raf. HA–RasV12 was cotransfected with GST–Raf in the presence or absence of HA–Sur-8. GST–Raf was purified by glutathione–agarose resin. The copurified Ras and HA–Sur-8 were detected by anti-HA for HA–Ras (second panel from top) and HA–Sur-8 (top) Western blots. The GST–Raf in the precipitation was detected by an anti-Raf Western blot (middle). Expressions of HA–Sur-8 and HA–RasV12 in cell lysates were shown in bottom two panels. (B) Sur-8C260Y and E457K mutants fail to enhance the interaction between Ras and Raf. Experiments are similar to A. (HA–Sur-8*) Sur-8C260Y in lanes 1–4 and Sur-8E457K in lanes 5–8, respectively. (C) Raf enhances Sur–8-Ras interaction in vitro. Cell lysates expressing HA–Sur-8 or Myc–Raf were mixed with purified GST–RasV12 protein (5 μg). GST–RasV12 and associated proteins were purified by glutathione–agarose and Western blotted with anti-HA to detect HA–Sur-8 (top) and anti-GST to detect GST–RasV12 (bottom). (Lane 6) A control of cell lysate containing HA–Sur-8. (D) Raf bridges the interaction between Ras and Sur-8ΔC. GST–Ras (5 μg) was incubated with cell lysates containing HA–Sur-8ΔC (containing residues 1–366) in the absence (lane 1) or presence (lane 2) of Raf containing cell lysates. HA–Sur-8ΔC (middle) and Raf (top) copurified by GST–Ras were detected by Western blot. GST–Ras coprecipitated HA–Sur-8ΔC only in the presence of Raf. (E) Sur-8, Ras, and Raf form a ternary complex. The 293 cells were transfected with various plasmids as indicated. Cell lysates were subjected to purification by glutathione–agarose beads. The purified proteins were eluted by glutathione and subjected to immunoprecipitation with anti-HA for HA–Ras followed by Western blot with anti-Raf (left). Similarly, proteins eluted from glutathione–agarose beads were immunoprecipitated with anti-Myc for Myc–Raf followed by Western blot with anti-Ras (middle). (Right) Experimental results using His–Sur-8 (lane 8) along with GST–Sur-8 (lane 9). His–Sur-8 complex was first purified by nickel agarose and eluted with imidazole. The eluted samples were immunoprecipitated with anti-Myc and followed by Western blot with anti-Raf.
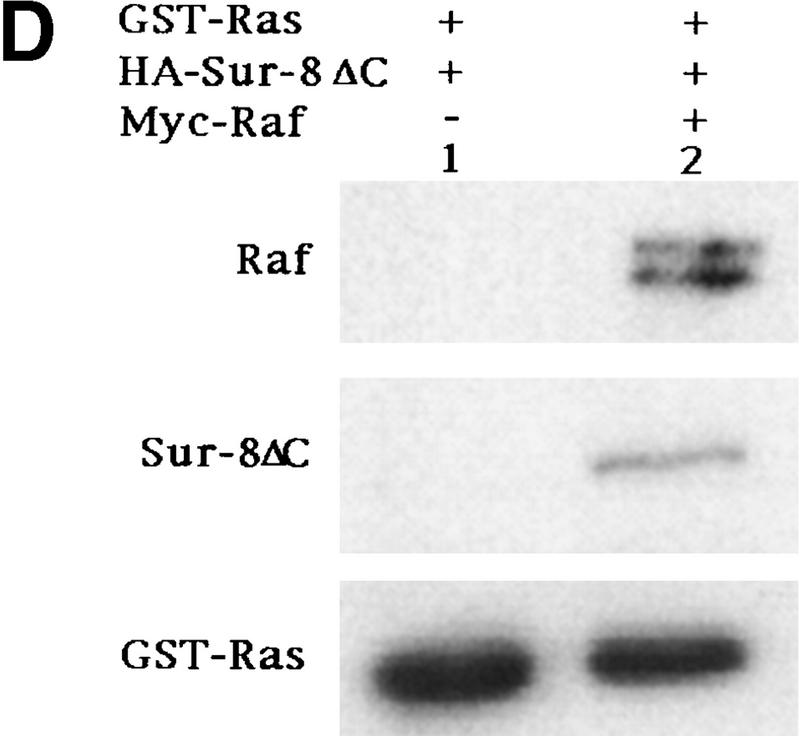
Sur-8 forms a complex with Ras and Raf. (A) Sur-8 enhances the interaction between Ras and Raf. HA–RasV12 was cotransfected with GST–Raf in the presence or absence of HA–Sur-8. GST–Raf was purified by glutathione–agarose resin. The copurified Ras and HA–Sur-8 were detected by anti-HA for HA–Ras (second panel from top) and HA–Sur-8 (top) Western blots. The GST–Raf in the precipitation was detected by an anti-Raf Western blot (middle). Expressions of HA–Sur-8 and HA–RasV12 in cell lysates were shown in bottom two panels. (B) Sur-8C260Y and E457K mutants fail to enhance the interaction between Ras and Raf. Experiments are similar to A. (HA–Sur-8*) Sur-8C260Y in lanes 1–4 and Sur-8E457K in lanes 5–8, respectively. (C) Raf enhances Sur–8-Ras interaction in vitro. Cell lysates expressing HA–Sur-8 or Myc–Raf were mixed with purified GST–RasV12 protein (5 μg). GST–RasV12 and associated proteins were purified by glutathione–agarose and Western blotted with anti-HA to detect HA–Sur-8 (top) and anti-GST to detect GST–RasV12 (bottom). (Lane 6) A control of cell lysate containing HA–Sur-8. (D) Raf bridges the interaction between Ras and Sur-8ΔC. GST–Ras (5 μg) was incubated with cell lysates containing HA–Sur-8ΔC (containing residues 1–366) in the absence (lane 1) or presence (lane 2) of Raf containing cell lysates. HA–Sur-8ΔC (middle) and Raf (top) copurified by GST–Ras were detected by Western blot. GST–Ras coprecipitated HA–Sur-8ΔC only in the presence of Raf. (E) Sur-8, Ras, and Raf form a ternary complex. The 293 cells were transfected with various plasmids as indicated. Cell lysates were subjected to purification by glutathione–agarose beads. The purified proteins were eluted by glutathione and subjected to immunoprecipitation with anti-HA for HA–Ras followed by Western blot with anti-Raf (left). Similarly, proteins eluted from glutathione–agarose beads were immunoprecipitated with anti-Myc for Myc–Raf followed by Western blot with anti-Ras (middle). (Right) Experimental results using His–Sur-8 (lane 8) along with GST–Sur-8 (lane 9). His–Sur-8 complex was first purified by nickel agarose and eluted with imidazole. The eluted samples were immunoprecipitated with anti-Myc and followed by Western blot with anti-Raf.
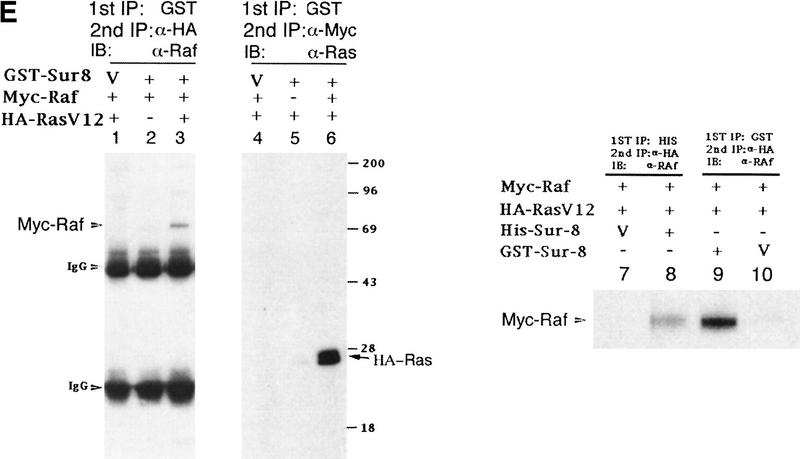
Sur-8 forms a complex with Ras and Raf. (A) Sur-8 enhances the interaction between Ras and Raf. HA–RasV12 was cotransfected with GST–Raf in the presence or absence of HA–Sur-8. GST–Raf was purified by glutathione–agarose resin. The copurified Ras and HA–Sur-8 were detected by anti-HA for HA–Ras (second panel from top) and HA–Sur-8 (top) Western blots. The GST–Raf in the precipitation was detected by an anti-Raf Western blot (middle). Expressions of HA–Sur-8 and HA–RasV12 in cell lysates were shown in bottom two panels. (B) Sur-8C260Y and E457K mutants fail to enhance the interaction between Ras and Raf. Experiments are similar to A. (HA–Sur-8*) Sur-8C260Y in lanes 1–4 and Sur-8E457K in lanes 5–8, respectively. (C) Raf enhances Sur–8-Ras interaction in vitro. Cell lysates expressing HA–Sur-8 or Myc–Raf were mixed with purified GST–RasV12 protein (5 μg). GST–RasV12 and associated proteins were purified by glutathione–agarose and Western blotted with anti-HA to detect HA–Sur-8 (top) and anti-GST to detect GST–RasV12 (bottom). (Lane 6) A control of cell lysate containing HA–Sur-8. (D) Raf bridges the interaction between Ras and Sur-8ΔC. GST–Ras (5 μg) was incubated with cell lysates containing HA–Sur-8ΔC (containing residues 1–366) in the absence (lane 1) or presence (lane 2) of Raf containing cell lysates. HA–Sur-8ΔC (middle) and Raf (top) copurified by GST–Ras were detected by Western blot. GST–Ras coprecipitated HA–Sur-8ΔC only in the presence of Raf. (E) Sur-8, Ras, and Raf form a ternary complex. The 293 cells were transfected with various plasmids as indicated. Cell lysates were subjected to purification by glutathione–agarose beads. The purified proteins were eluted by glutathione and subjected to immunoprecipitation with anti-HA for HA–Ras followed by Western blot with anti-Raf (left). Similarly, proteins eluted from glutathione–agarose beads were immunoprecipitated with anti-Myc for Myc–Raf followed by Western blot with anti-Ras (middle). (Right) Experimental results using His–Sur-8 (lane 8) along with GST–Sur-8 (lane 9). His–Sur-8 complex was first purified by nickel agarose and eluted with imidazole. The eluted samples were immunoprecipitated with anti-Myc and followed by Western blot with anti-Raf.
References
-
- Chang HC, Karim FD, O'Neill EM, Rebay I, Solomon NM, Therrien M, Wassarman DA, Wolff T, Rubin GM. Ras signal transduction pathway in Drosophila eye development. Cold Spring Harb Symp Quant Biol. 1994;59:147–153. - PubMed
-
- Derijard B, Hibi M, Wu IH, Barrett T, Su B, Deng T, Karin M, Davis RJ. JNK1: A protein kinase stimulated by UV light and Ha-Ras that binds and phosphorylates the c-Jun activation domain. Cell. 1994;76:1025–1037. - PubMed
-
- Field J, Xu HP, Michaeli T, Ballester R, Sass P, Wigler M, Colicelli J. Mutations of the adenylyl cyclase gene that block RAS function in Saccharomyces cerevisiae. Science. 1990;247:464–467. - PubMed
-
- Guan KL, Dixon JE. Eukaryotic proteins expressed in Escherichia coli: An improved thrombin cleavage and purification procedure of fusion proteins with glutathione S-transferase. Anal Biochem. 1991;192:262–267. - PubMed
-
- Howe LR, Leevers SJ, Gomez N, Nakielny S, Cohen P, Marshall CJ. Activation of the MAP kinase pathway by the protein kinase raf. Cell. 1992;71:335–342. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous