

Epsin 1 undergoes nucleocytosolic shuttling and its eps15 interactor NH(2)-terminal homology (ENTH) domain, structurally similar to Armadillo and HEAT repeats, interacts with the transcription factor promyelocytic leukemia Zn(2)+ finger protein (PLZF)
- PMID: 10791968
- PMCID: PMC2174850
- DOI: 10.1083/jcb.149.3.537
Epsin 1 undergoes nucleocytosolic shuttling and its eps15 interactor NH(2)-terminal homology (ENTH) domain, structurally similar to Armadillo and HEAT repeats, interacts with the transcription factor promyelocytic leukemia Zn(2)+ finger protein (PLZF)
Abstract
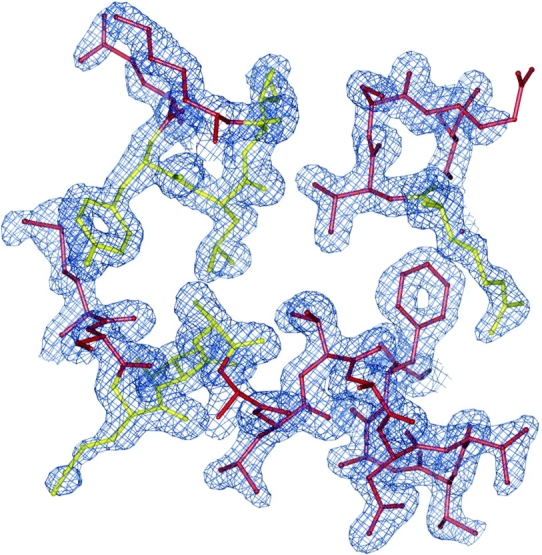
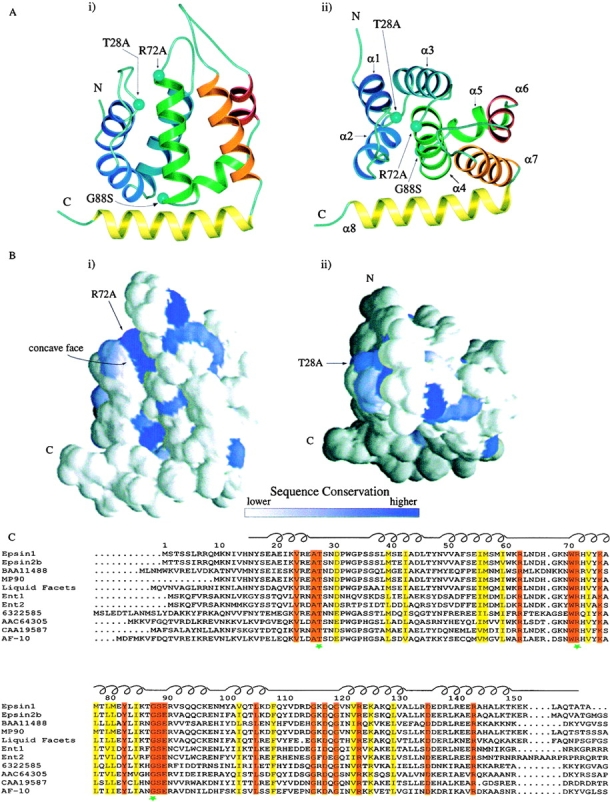
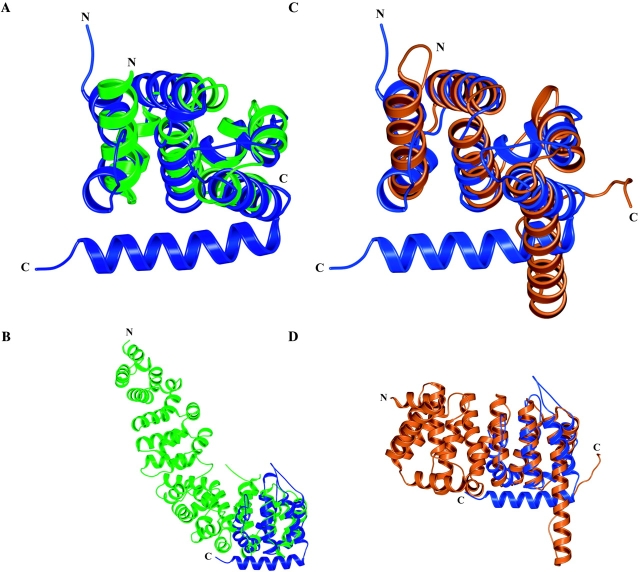
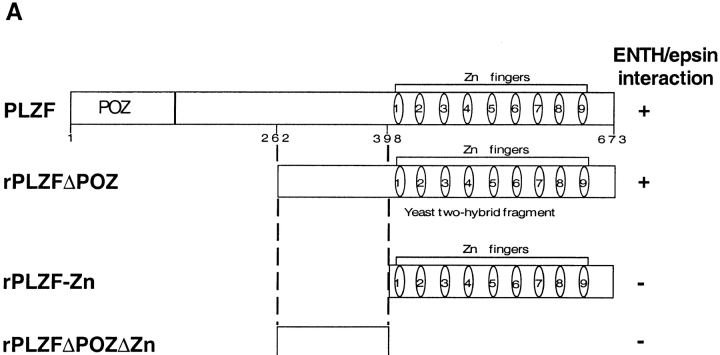
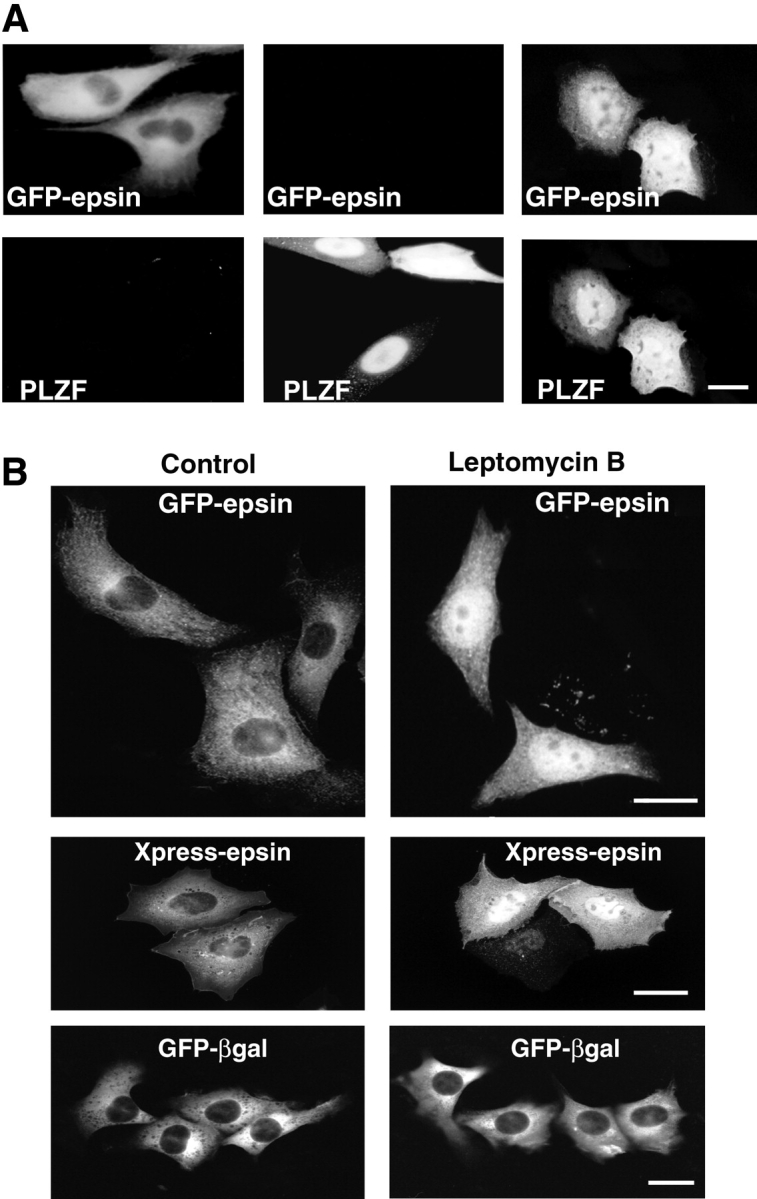
Epsin (Eps15 interactor) is a cytosolic protein involved in clathrin-mediated endocytosis via its direct interactions with clathrin, the clathrin adaptor AP-2, and Eps15. The NH(2)-terminal portion of epsin contains a phylogenetically conserved module of unknown function, known as the ENTH domain (epsin NH(2)-terminal homology domain). We have now solved the crystal structure of rat epsin 1 ENTH domain to 1.8 A resolution. This domain is structurally similar to armadillo and Heat repeats of beta-catenin and karyopherin-beta, respectively. We have also identified and characterized the interaction of epsin 1, via the ENTH domain, with the transcription factor promyelocytic leukemia Zn(2)+ finger protein (PLZF). Leptomycin B, an antifungal antibiotic, which inhibits the Crm1- dependent nuclear export pathway, induces an accumulation of epsin 1 in the nucleus. These findings suggest that epsin 1 may function in a signaling pathway connecting the endocytic machinery to the regulation of nuclear function.
Figures

References
-
- Abrahams J.P., Leslie A.G.W. Methods used in the structure determination of bovine mitochondrial F1 ATPase. Acta Crystallogr. 1996;D52:30–42. - PubMed
-
- Barth A.I.M., Näthke I.S., Nelson W.J. Cadherins, catenins and APC proteininterplay between cytoskeletal complexes and signalling pathways. Curr. Opin. Cell Biol. 1997;9:683–690. - PubMed
-
- Bricogne G. Bayesian statistical viewpoint on structure determinationbasic concepts and examples. Methods Enzymol. 1997;276:361–423. - PubMed
-
- Brunger A.T. Free R valuea novel statistical quantity for assessing the accuracy of crystal structures. Nature. 1992;355:472–475. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
