

Epidermal growth factor receptor expression in neurofibromatosis type 1-related tumors and NF1 animal models
- PMID: 10791998
- PMCID: PMC315438
- DOI: 10.1172/JCI7610
Epidermal growth factor receptor expression in neurofibromatosis type 1-related tumors and NF1 animal models
Abstract
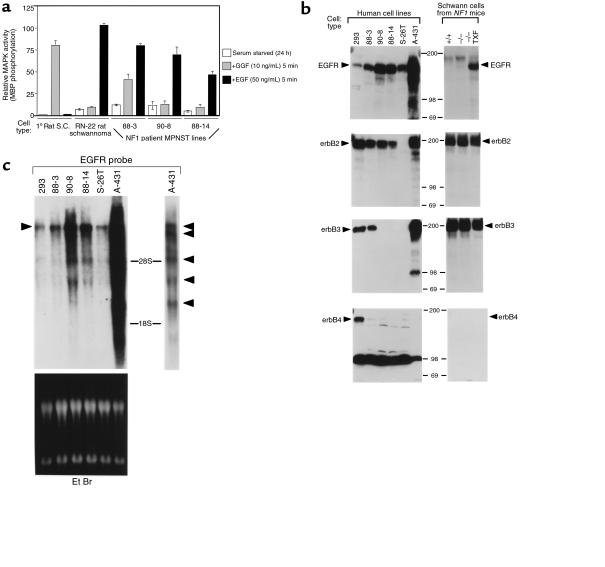
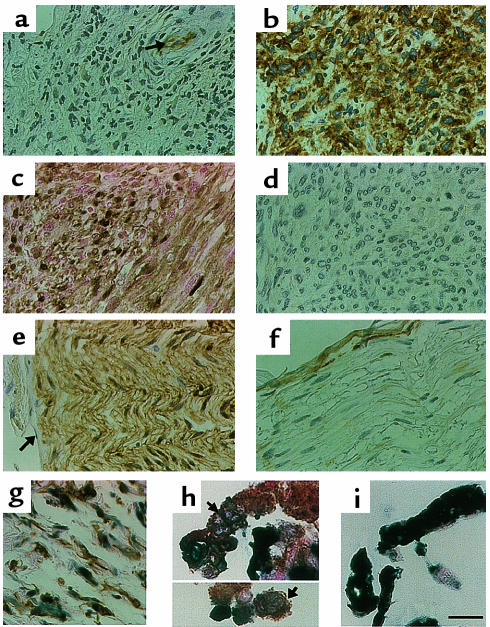
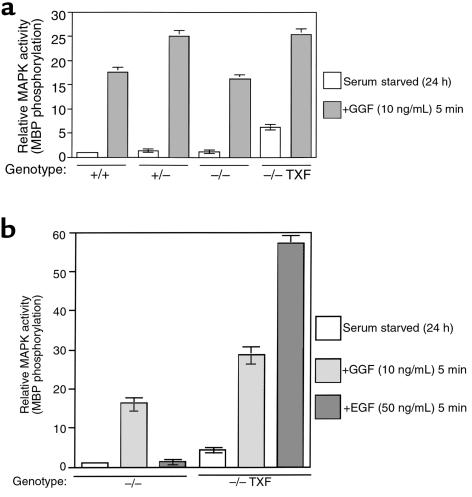
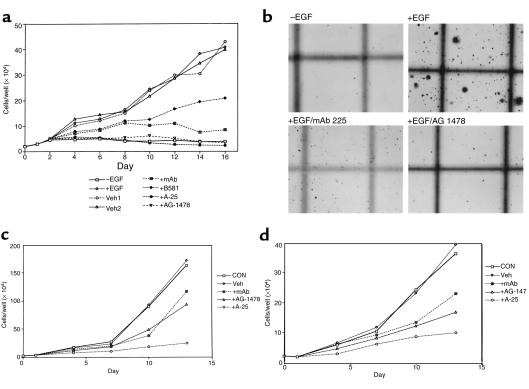
We have found that EGF-R expression is associated with the development of the Schwann cell-derived tumors characteristic of neurofibromatosis type 1 (NF1) and in animal models of this disease. This is surprising, because Schwann cells normally lack EGF-R and respond to ligands other than EGF. Nevertheless, immunoblotting, Northern analysis, and immunohistochemistry revealed that each of 3 malignant peripheral nerve sheath tumor (MPNST) cell lines from NF1 patients expressed the EGF-R, as did 7 of 7 other primary MPNSTs, a non-NF1 MPNST cell line, and the S100(+) cells from each of 9 benign neurofibromas. Furthermore, transformed derivatives of Schwann cells from NF1(-/-) mouse embryos also expressed the EGF-R. All of the cells or cell lines expressing EGF-R responded to EGF by activation of downstream signaling pathways. Thus, EGF-R expression may play an important role in NF1 tumorigenesis and Schwann cell transformation. Consistent with this hypothesis, growth of NF1 MPNST lines and the transformed NF1(-/-) mouse embryo Schwann cells was greatly stimulated by EGF in vitro and could be blocked by agents that antagonize EGF-R function.
Figures
References
-
- Huson SM, Harper PS, Compston D. Von Recklinghausen neurofibromatosis: a clinical and population study in south-east Wales. Brain. 1988;111:1355–1381. - PubMed
-
- Upadhyaya, M., and Cooper, D.N. 1998. Neurofibromatosis type 1: from genotype to phenotype. BIOS Scientific Publishers Ltd. Oxford, United Kingdom. pp. 230.
-
- Ferner, R.E. 1998. Clinical aspects of neurofibromatosis 1. In Neurofibromatosis type 1: from genotype to phenotype. M. Upadhyaya and D.N. Cooper, editors. BIOS Scientific Publishers Ltd., Oxford, United Kingdom. 21–38.
-
- Rosenbaum T, Petrie KM, Ratner N. Neurofibromatosis type 1: genetic and cellular mechanisms of peripheral nerve tumor formation. The Neuroscientist. 1997;3:412–420.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
