

Corticotropin-releasing hormone links pituitary adrenocorticotropin gene expression and release during adrenal insufficiency
- PMID: 10792002
- PMCID: PMC315436
- DOI: 10.1172/JCI5250
Corticotropin-releasing hormone links pituitary adrenocorticotropin gene expression and release during adrenal insufficiency
Abstract
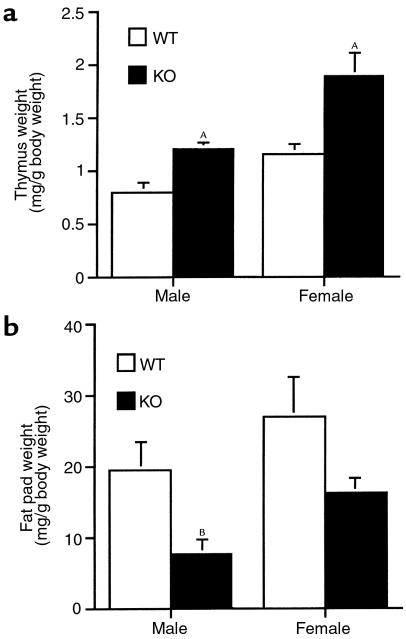
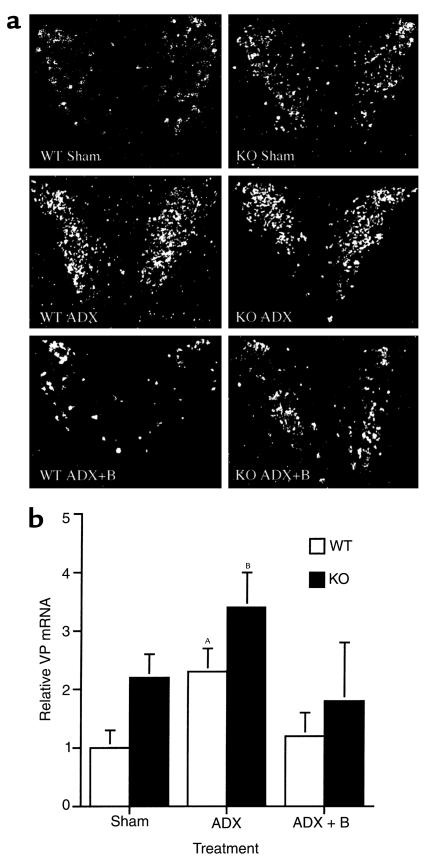
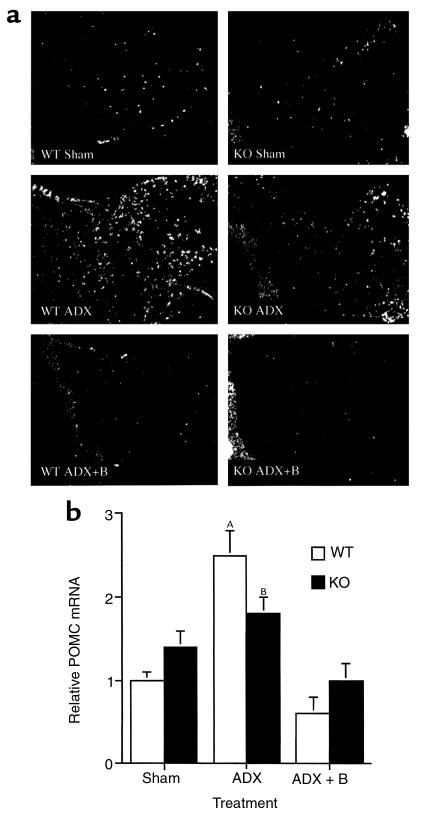
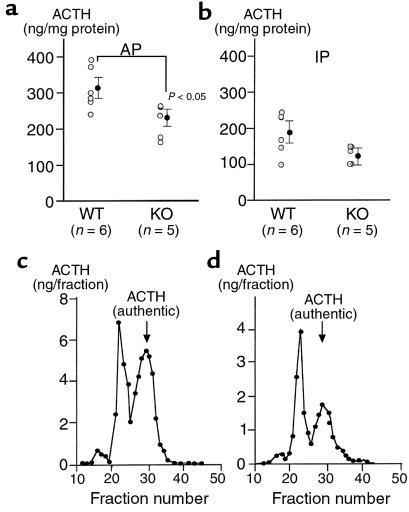
Corticotropin-releasing hormone (CRH)-deficient (KO) mice provide a unique system to define the role of CRH in regulation of the hypothalamic-pituitary-adrenal (HPA) axis. Despite several manifestations of chronic glucocorticoid insufficiency, basal pituitary proopiomelanocortin (POMC) mRNA, adrenocorticotrophic hormone (ACTH) peptide content within the pituitary, and plasma ACTH concentrations are not elevated in CRH KO mice. The normal POMC mRNA content in KO mice is dependent upon residual glucocorticoid secretion, as it increases in both KO and WT mice after adrenalectomy; this increase is reversed by glucocorticoid, but not aldosterone, replacement. However, the normal plasma levels of ACTH in CRH KO mice are not dependent upon residual glucocorticoid secretion, because, after adrenalectomy, these levels do not undergo the normal increase seen in KO mice despite the increase in POMC mRNA content. Administration of CRH restores ACTH secretion to its expected high level in adrenalectomized CRH KO mice. Thus, in adrenal insufficiency, loss of glucocorticoid feedback by itself can increase POMC gene expression in the pituitary; but CRH action is essential for this to result in increased secretion of ACTH. This may explain why, after withdrawal of chronic glucocorticoid treatment, reactivation of CRH secretion is a necessary prerequisite for recovery from suppression of the HPA axis.
Figures
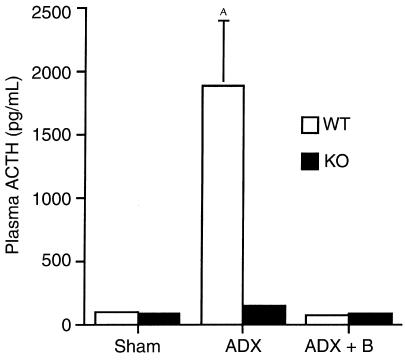
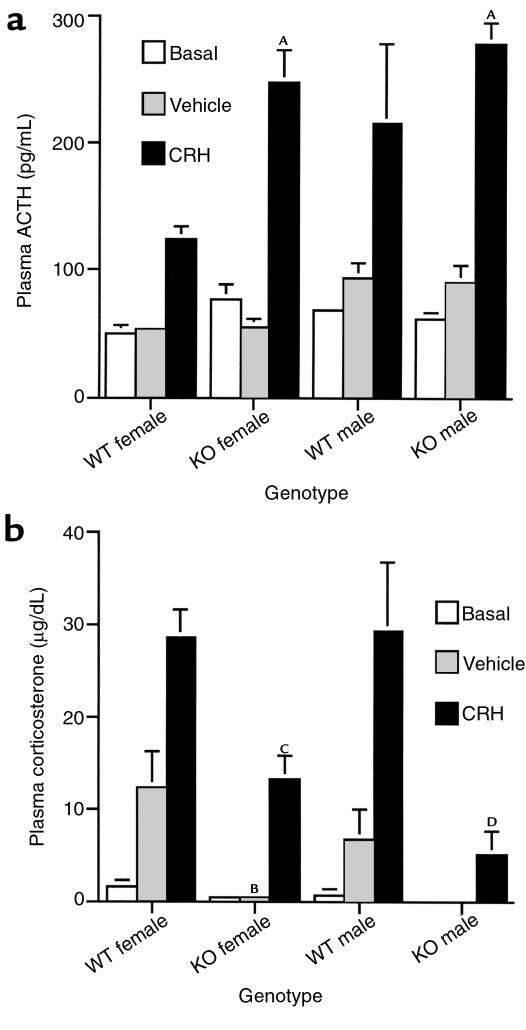
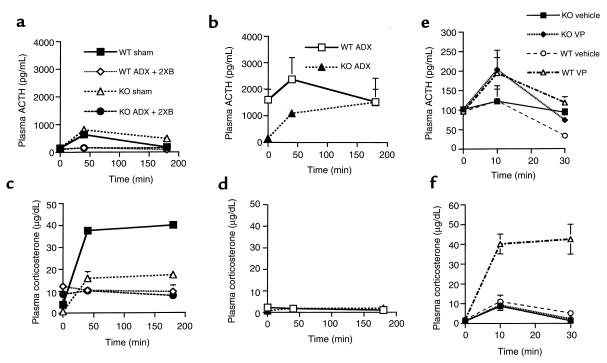
Comment in
-
Regulation of adrenocorticotropic hormone secretion: lessons from mice deficient in corticotropin-releasing hormone.J Clin Invest. 2000 May;105(9):1187-8. doi: 10.1172/JCI10002. J Clin Invest. 2000. PMID: 10791992 Free PMC article. No abstract available.
References
-
- Chrousos GP. The hypothalamic-pituitary-adrenal axis and immune-mediated inflammation. N Engl J Med. 1995;332:1351–1362. - PubMed
-
- Munck A, Guyre PM, Holbrook NJ. Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocr Rev. 1984;5:25–44. - PubMed
-
- Orth, D.N., Kovacs, W.J., and Debold, C.R. 1992. The adrenal cortex. In Williams textbook of endocrinology. J.D. Wilson and D.W. Foster, editors. W.B. Saunders Co. Philadelphia, Pennsylvania, USA. 489–619.
-
- Suda T, et al. The role of corticotropin-releasing factor and vasopressin in hypoglycemia induced proopiomelanocortin gene expression in the rat anterior pituitary gland. Brain Res. 1992;579:303–308. - PubMed
-
- Plotsky PM, Bruhn TO, Vale W. Hypophysiotropic regulation of adrenocorticotropin secretion in response to insulin-induced hypoglycemia. Endocrinology. 1985;117:323–329. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
