

Hsp90 is essential for the synthesis and subsequent membrane association, but not the maintenance, of the Src-kinase p56(lck)
- PMID: 10793137
- PMCID: PMC14869
- DOI: 10.1091/mbc.11.5.1585
Hsp90 is essential for the synthesis and subsequent membrane association, but not the maintenance, of the Src-kinase p56(lck)
Abstract
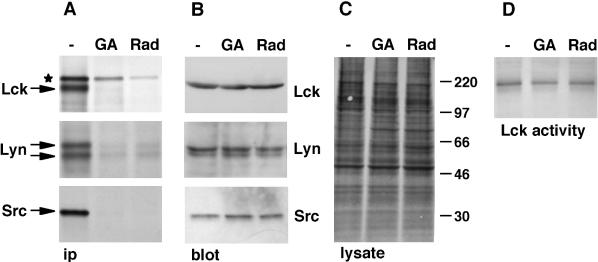
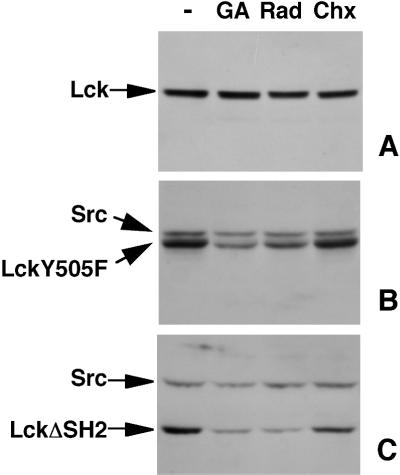
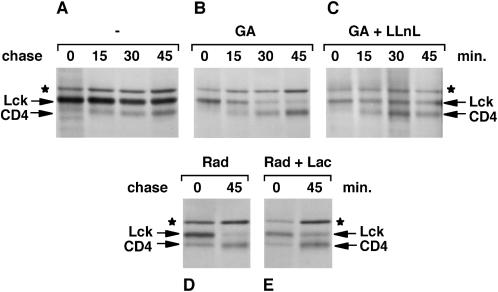
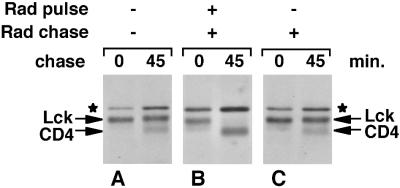
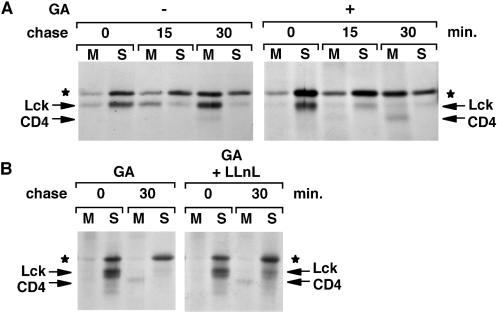
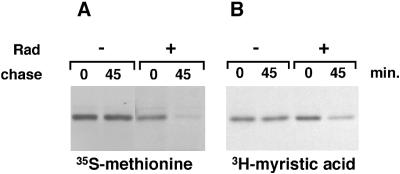
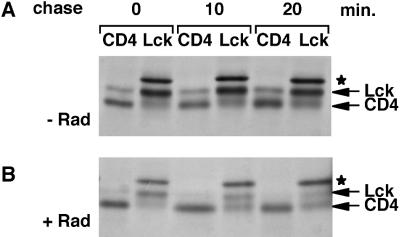
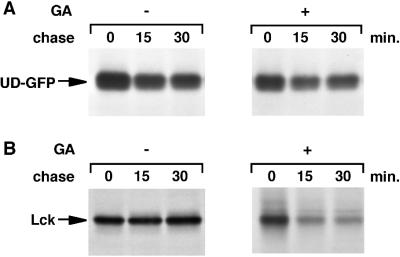
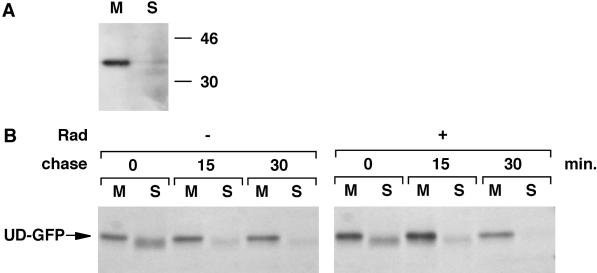
Tyrosine kinases of the Src family are synthesized as cytosolic proteins that subsequently translocate to membranes. Little is known of the mechanisms responsible for targeting these proteins to membranes, although a role for the cytosolic chaperone Hsp90 has been proposed. Here, we have studied the involvement of Hsp90 in the synthesis, membrane binding, and maintenance of the Src-kinase Lck. Using specific inhibitors of Hsp90, geldanamycin and radicicol, we found that functional Hsp90 is essential for the stability of newly synthesized, but not mature, Lck. Similar results were obtained for two other Src-kinases, c-Src and Lyn. In contrast, LckY505F and LckDeltaSH2, constitutively active Lck mutants lacking the C-terminal regulatory tyrosine or the entire Src homology 2 domain, respectively, required Hsp90 activity to stabilize the mature proteins. Lck synthesized in the absence of Hsp90 activity was degraded within 30-45 min. This unstable Lck was myristoylated normally but did not associate with membranes or CD4, interactions that normally start within minutes of the completion of Lck synthesis. A construct composed of the N-terminal unique domain of Lck fused to green fluorescent protein did not require Hsp90 activity during synthesis. In addition, this protein associated with membranes efficiently in the absence of Hsp90 activity. Together these data suggest that interaction with Hsp90 is necessary for the correct synthesis and subsequent membrane binding of Lck. However, Hsp90 does not appear to play a direct role in Lck membrane, or CD4, association.
Figures
References
-
- Bhatnagar RS, Gordon JI. Understanding covalent modifications of proteins by lipids: where cell biology and biophysics mingle. Trends Cell Biol. 1997;7:14–21. - PubMed
-
- Brouns GS, de Vries E, van Noesel CJ, Mason DY, van Lier RA, Borst J. The structure of the mu/pseudo light chain complex on human pre-B cells is consistent with a function in signal transduction. Eur J Immunol. 1993;23:1088–1097. - PubMed
-
- Brown MT, Cooper JA. Regulation, substrates and functions of src. Biochim Biophys Acta. 1996;1287:121–149. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
