

Influenza A viruses lacking sialidase activity can undergo multiple cycles of replication in cell culture, eggs, or mice
- PMID: 10799596
- PMCID: PMC110874
- DOI: 10.1128/jvi.74.11.5206-5212.2000
Influenza A viruses lacking sialidase activity can undergo multiple cycles of replication in cell culture, eggs, or mice
Abstract
Influenza A viruses possess both hemagglutinin (HA), which is responsible for binding to the terminal sialic acid of sialyloligosaccharides on the cell surface, and neuraminidase (NA), which contains sialidase activity that removes sialic acid from sialyloligosaccharides. Interplay between HA receptor-binding and NA receptor-destroying sialidase activity appears to be important for replication of the virus. Previous studies by others have shown that influenza A viruses lacking sialidase activity can undergo multiple cycles of replication if sialidase activity is provided exogenously. To investigate the sialidase requirement of influenza viruses further, we generated a series of sialidase-deficient mutants. Although their growth was less efficient than that of the parental NA-dependent virus, these viruses underwent multiple cycles of replication in cell culture, eggs, and mice. To understand the molecular basis of this viral growth adaptation in the absence of sialidase activity, we investigated changes in the HA receptor-binding affinity of the sialidase-deficient mutants. The results show that mutations around the HA receptor-binding pocket reduce the virus's affinity for cellular receptors, compensating for the loss of sialidase. Thus, sialidase activity is not absolutely required in the influenza A virus life cycle but appears to be necessary for efficient virus replication.
Figures
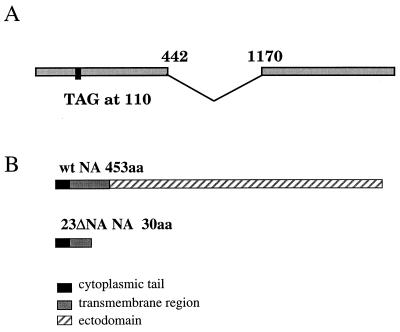
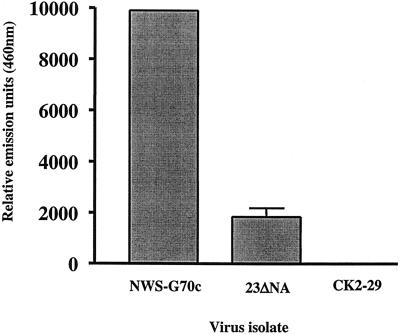
References
-
- Air G M, Laver W G. The neuraminidase of influenza virus. Proteins Struct Funct Genet. 1989;6:341–356. - PubMed
-
- Blick T J, Sahasrabudhe A, McDonald M, Owens I J, Morley P J, Fenton R J, McKimm-Breschkin J L. The interaction of neuraminidase and hemagglutinin mutations in influenza virus resistance to 4-guanidino-Neu5Ac2en. Virology. 1998;246:95–103. - PubMed
-
- Callan R J, Early G, Kida H, Hinshaw V S. The appearance of H3 influenza viruses in seals. J Gen Virol. 1995;76:199–203. - PubMed
-
- Crecelius D M, Deom C M, Schulze I T. Biological properties of a hemagglutinin mutant of influenza virus selected by host cells. Virology. 1984;139:164–177. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
