

hnRNP C is required for postimplantation mouse development but Is dispensable for cell viability
- PMID: 10805751
- PMCID: PMC85779
- DOI: 10.1128/MCB.20.11.4094-4105.2000
hnRNP C is required for postimplantation mouse development but Is dispensable for cell viability
Abstract
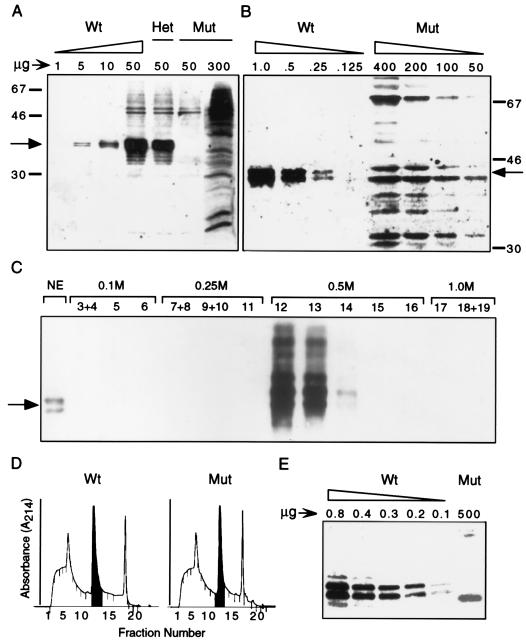
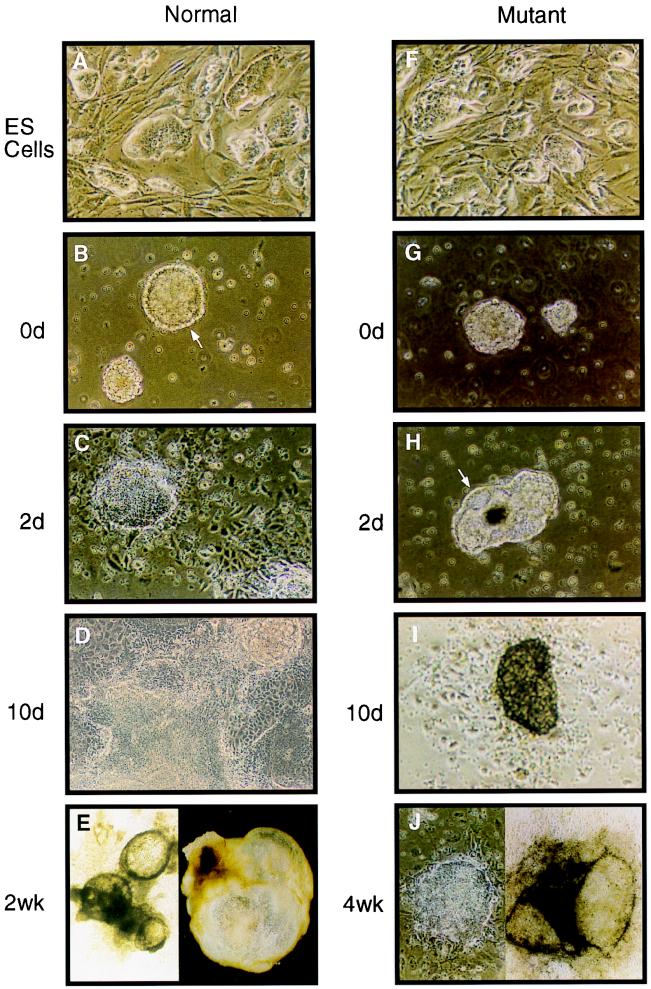
The hnRNP C1 and C2 proteins are among the most abundant proteins in the nucleus, and as ubiquitous components of RNP complexes, they have been implicated in many aspects of mRNA biogenesis. In this report, we have characterized a null mutation induced in embryonic stem cells by insertion of the U3His gene trap retrovirus into the first intron of the hnRNP C1/C2 gene. cDNAs encoding murine hnRNP C1 and C2 were characterized, and the predicted protein sequences were found to be highly conserved among vertebrates. A human consensus sequence, generated from over 400 expressed sequence tags, suggests two revisions to the previously published human sequence. In addition, alternatively spliced transcripts, expressed only by the murine gene, encode four novel proteins: variants of C1 and C2 with either seven additional amino acids or one fewer amino acid in a region between the oligomerization and C-terminal acidic domains. The disrupted gene was transmitted into the germ line and is tightly linked to a recessive, embryonic lethal phenotype. Homozygous mutant embryos fail to develop beyond the egg cylinder stage and are resorbed by 10.5 days of gestation, a phenotype consistent with a fundamental role in cellular metabolism. However, hnRNP C1 and C2 are not required for cell viability. Embryonic stem cell lines established from homozygous mutant blastocysts did not express detectable levels of either protein yet were able to grow and differentiate in vitro, albeit more slowly than wild-type cells. These results indicate that the C1 and C2 hnRNPs are not required for any essential step in mRNA biogenesis; however, the proteins may influence the rate and/or fidelity of one or more steps.
Figures

Similar articles
-
Primary structure of human nuclear ribonucleoprotein particle C proteins: conservation of sequence and domain structures in heterogeneous nuclear RNA, mRNA, and pre-rRNA-binding proteins.Mol Cell Biol. 1987 May;7(5):1731-9. doi: 10.1128/mcb.7.5.1731-1739.1987. Mol Cell Biol. 1987. PMID: 3110598 Free PMC article.
-
The determinants of RNA-binding specificity of the heterogeneous nuclear ribonucleoprotein C proteins.J Biol Chem. 1994 Sep 16;269(37):23074-8. J Biol Chem. 1994. PMID: 8083209
-
Arginine N-methyltransferase 1 is required for early postimplantation mouse development, but cells deficient in the enzyme are viable.Mol Cell Biol. 2000 Jul;20(13):4859-69. doi: 10.1128/MCB.20.13.4859-4869.2000. Mol Cell Biol. 2000. PMID: 10848611 Free PMC article.
-
A model for heterogeneous nuclear ribonucleoproteins in telomere and telomerase regulation.Oncogene. 2002 Jan 21;21(4):580-3. doi: 10.1038/sj.onc.1205086. Oncogene. 2002. PMID: 11850782 Review.
-
Functions of heterogeneous nuclear ribonucleoproteins in stem cell potency and differentiation.Biomed Res Int. 2013;2013:623978. doi: 10.1155/2013/623978. Epub 2013 Jul 29. Biomed Res Int. 2013. PMID: 23984388 Free PMC article. Review.
Cited by
-
Differential alternative splicing landscape identifies potentially functional RNA binding proteins in early embryonic development in mammals.iScience. 2024 Feb 2;27(3):109104. doi: 10.1016/j.isci.2024.109104. eCollection 2024 Mar 15. iScience. 2024. PMID: 38433915 Free PMC article.
-
Multiomics profiling uncovers curdione-induced reproductive toxicity in HTR-8/SVneo cells.Heliyon. 2024 Sep 27;10(21):e38650. doi: 10.1016/j.heliyon.2024.e38650. eCollection 2024 Nov 15. Heliyon. 2024. PMID: 39524727 Free PMC article.
-
Regulation of protein kinase CK1alphaLS by dephosphorylation in response to hydrogen peroxide.Arch Biochem Biophys. 2007 Oct 15;466(2):242-9. doi: 10.1016/j.abb.2007.06.010. Epub 2007 Jun 21. Arch Biochem Biophys. 2007. PMID: 17626781 Free PMC article.
-
PLRG1 is an essential regulator of cell proliferation and apoptosis during vertebrate development and tissue homeostasis.Mol Cell Biol. 2009 Jun;29(11):3173-85. doi: 10.1128/MCB.01807-08. Epub 2009 Mar 23. Mol Cell Biol. 2009. PMID: 19307306 Free PMC article.
-
DAZAP1, an hnRNP protein, is required for normal growth and spermatogenesis in mice.RNA. 2008 Sep;14(9):1814-22. doi: 10.1261/rna.1152808. Epub 2008 Jul 30. RNA. 2008. PMID: 18669443 Free PMC article.
References
-
- Bachvarova R, De Leon V. Polyadenylated RNA of mouse ova and loss of maternal RNA in early development. Dev Biol. 1980;74:1–8. - PubMed
-
- Barnett S F, LeStourgeon W M, Friedman D L. Rapid purification of native C protein from nuclear ribonucleoprotein particles. J Biochem Biophys Methods. 1988;16:87–97. - PubMed
-
- Bennett M, Michaud S, Kingston J, Reed R. Protein components specifically associated with prespliceosome and spliceosome complexes. Genes Dev. 1992;6:1986–2000. - PubMed
-
- Beyer A L, Christensen M E, Walker B W, LeStourgeon W M. Identification and characterization of the packaging proteins of core 40S hnRNP particles. Cell. 1977;11:127–138. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous