

S-Phase progression mediates activation of a silenced gene in synthetic nuclei
- PMID: 10805758
- PMCID: PMC85786
- DOI: 10.1128/MCB.20.11.4169-4180.2000
S-Phase progression mediates activation of a silenced gene in synthetic nuclei
Abstract
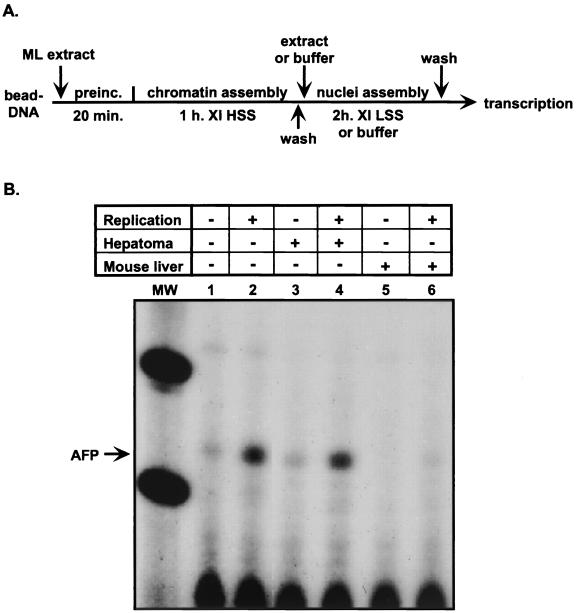
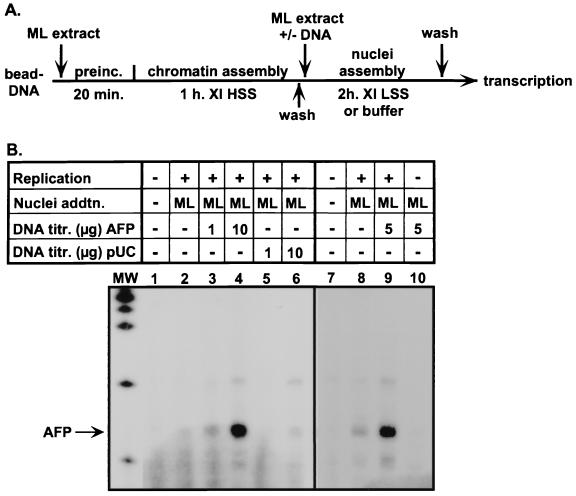
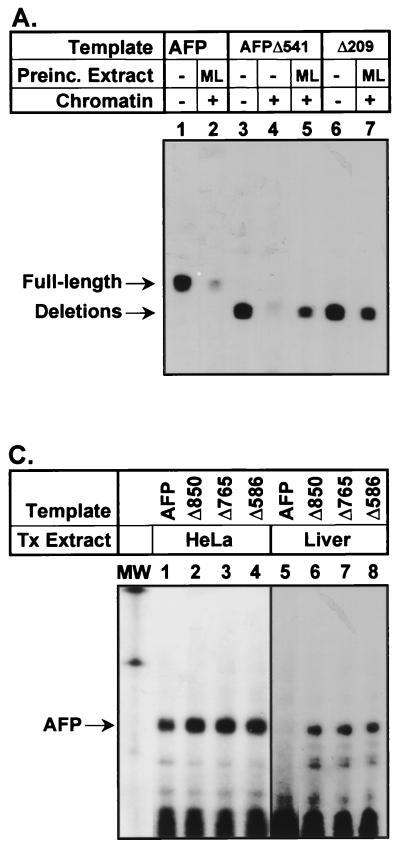
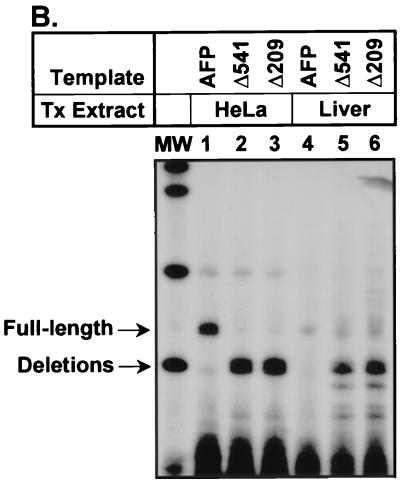
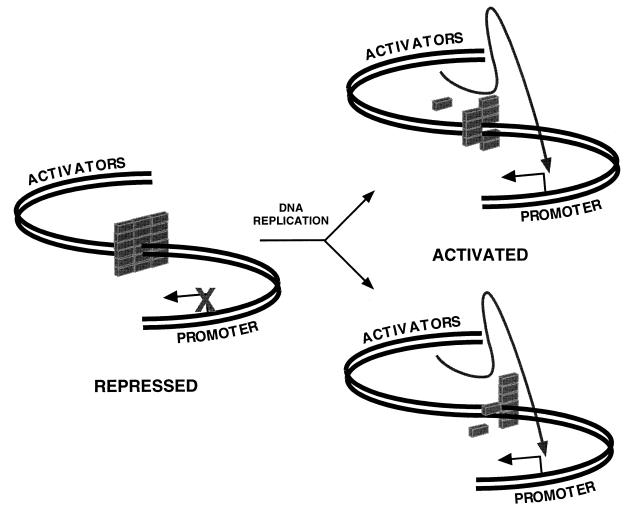
Aberrant expression of developmentally silenced genes, characteristic of tumor cells and regenerating tissue, is highly correlated with increased cell proliferation. By modeling this process in vitro in synthetic nuclei, we find that DNA replication leads to deregulation of established developmental expression patterns. Chromatin assembly in the presence of adult mouse liver nuclear extract mediates developmental stage-specific silencing of the tumor marker gene alpha-fetoprotein (AFP). Replication of silenced AFP chromatin in synthetic nuclei depletes sequence-specific transcription repressors, thereby disrupting developmentally regulated repression. Hepatoma-derived factors can target partial derepression of AFP, but full transcription activation requires DNA replication. Thus, unscheduled entry into S phase directly mediates activation of a developmentally silenced gene by (i) depleting developmental stage-specific transcription repressors and (ii) facilitating binding of transactivators.
Figures
Similar articles
-
Hepatocyte nuclear factor 3 relieves chromatin-mediated repression of the alpha-fetoprotein gene.J Biol Chem. 1999 Aug 27;274(35):25113-20. doi: 10.1074/jbc.274.35.25113. J Biol Chem. 1999. PMID: 10455192
-
Transcription factor interactions and chromatin modifications associated with p53-mediated, developmental repression of the alpha-fetoprotein gene.Mol Cell Biol. 2005 Mar;25(6):2147-57. doi: 10.1128/MCB.25.6.2147-2157.2005. Mol Cell Biol. 2005. PMID: 15743813 Free PMC article.
-
Functional analysis of developmentally regulated chromatin-hypersensitive domains carrying the alpha 1-fetoprotein gene promoter and the albumin/alpha 1-fetoprotein intergenic enhancer.Mol Cell Biol. 1993 Mar;13(3):1619-33. doi: 10.1128/mcb.13.3.1619-1633.1993. Mol Cell Biol. 1993. PMID: 7680097 Free PMC article.
-
p53-mediated repression of alpha-fetoprotein gene expression by specific DNA binding.Mol Cell Biol. 1999 Feb;19(2):1279-88. doi: 10.1128/MCB.19.2.1279. Mol Cell Biol. 1999. PMID: 9891062 Free PMC article.
-
[Several transcription factors participate in the functioning of the alpha-fetoprotein gene promoter].Bull Cancer. 1995 Jul;82(7):541-50. Bull Cancer. 1995. PMID: 7549116 Review. French.
Cited by
-
p53-targeted LSD1 functions in repression of chromatin structure and transcription in vivo.Mol Cell Biol. 2008 Sep;28(17):5139-46. doi: 10.1128/MCB.00287-08. Epub 2008 Jun 23. Mol Cell Biol. 2008. PMID: 18573881 Free PMC article.
-
De novo assembly of genuine replication forks on an immobilized circular plasmid in Xenopus egg extracts.Nucleic Acids Res. 2006 Jul 26;34(13):e91. doi: 10.1093/nar/gkl512. Nucleic Acids Res. 2006. PMID: 16870720 Free PMC article.
-
Vertebrate HoxB gene expression requires DNA replication.EMBO J. 2003 Jul 15;22(14):3737-48. doi: 10.1093/emboj/cdg352. EMBO J. 2003. PMID: 12853488 Free PMC article.
-
A direct intersection between p53 and transforming growth factor beta pathways targets chromatin modification and transcription repression of the alpha-fetoprotein gene.Mol Cell Biol. 2005 Feb;25(3):1200-12. doi: 10.1128/MCB.25.3.1200-1212.2005. Mol Cell Biol. 2005. PMID: 15657445 Free PMC article.
-
Zinc finger protein ZBTB20 is a key repressor of alpha-fetoprotein gene transcription in liver.Proc Natl Acad Sci U S A. 2008 Aug 5;105(31):10859-64. doi: 10.1073/pnas.0800647105. Epub 2008 Jul 31. Proc Natl Acad Sci U S A. 2008. PMID: 18669658 Free PMC article.
References
-
- Adams C C, Workman J L. Nucleosome displacement in transcription. Cell. 1993;72:305–308. - PubMed
-
- Aparicio O M, Gottschling D E. Overcoming telomeric silencing: a trans-activator competes to establish gene expression in a cell cycle-dependent way. Genes Dev. 1994;8:1133–1146. - PubMed
-
- Armstrong J A, Bieker J J, Emerson B M. A SWI/SNF-related chromatin remodeling complex, E-RC1, is required for tissue-specific transcriptional regulation by EKLF in vitro. Cell. 1998;95:93–104. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current protocols in molecular biology. Boston, Mass: Current Protocols; 1987.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources