

Rapid evolution in plant chitinases: molecular targets of selection in plant-pathogen coevolution
- PMID: 10805791
- PMCID: PMC25827
- DOI: 10.1073/pnas.97.10.5322
Rapid evolution in plant chitinases: molecular targets of selection in plant-pathogen coevolution
Abstract
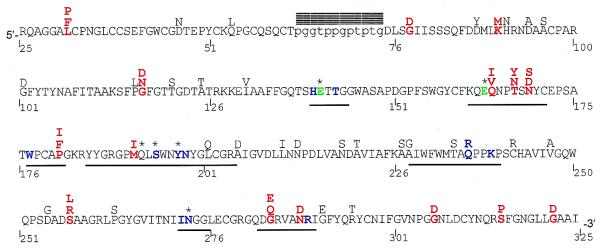
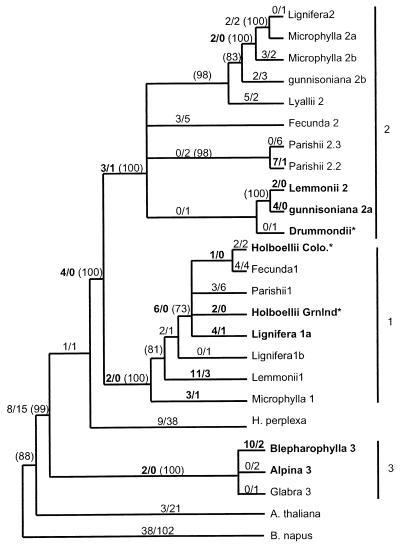
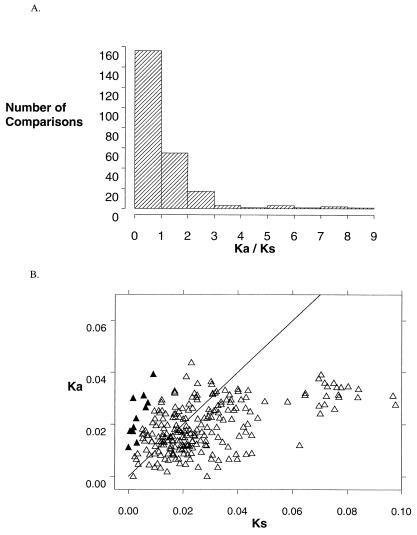
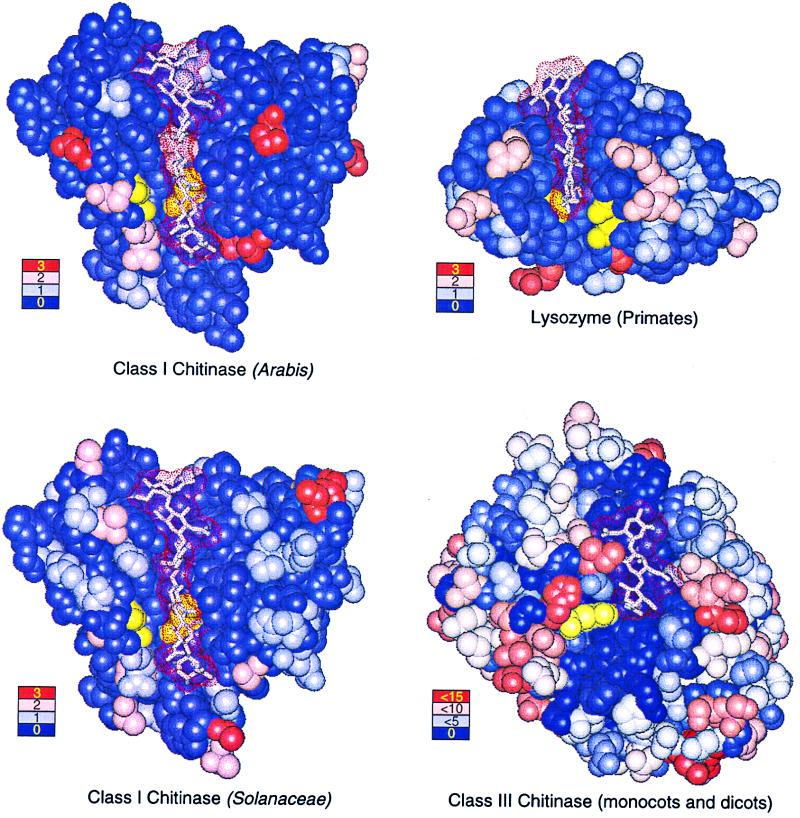
Many pathogen recognition genes, such as plant R-genes, undergo rapid adaptive evolution, providing evidence that these genes play a critical role in plant-pathogen coevolution. Surprisingly, whether rapid adaptive evolution also occurs in genes encoding other kinds of plant defense proteins is unknown. Unlike recognition proteins, plant chitinases attack pathogens directly, conferring disease resistance by degrading chitin, a component of fungal cell walls. Here, we show that nonsynonymous substitution rates in plant class I chitinase often exceed synonymous rates in the plant genus Arabis (Cruciferae) and in other dicots, indicating a succession of adaptively driven amino acid replacements. We identify individual residues that are likely subject to positive selection by using codon substitution models and determine the location of these residues on the three-dimensional structure of class I chitinase. In contrast to primate lysozymes and plant class III chitinases, structural and functional relatives of class I chitinase, the adaptive replacements of class I chitinase occur disproportionately in the active site cleft. This highly unusual pattern of replacements suggests that fungi directly defend against chitinolytic activity through enzymatic inhibition or other forms of chemical resistance and identifies target residues for manipulating chitinolytic activity. These data also provide empirical evidence that plant defense proteins not involved in pathogen recognition also evolve in a manner consistent with rapid coevolutionary interactions.
Figures
References
-
- Thompson J N, Burdon J J. Nature (London) 1992;360:121–125.
-
- Simms E L. BioScience. 1996;46:136–145.
-
- Stahl E A, Dwyer G, Mauricio R, Kreitman M, Bergelson J. Nature (London) 1999;400:667–671. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
