

Antibody repertoire development in fetal and newborn piglets, III. Colonization of the gastrointestinal tract selectively diversifies the preimmune repertoire in mucosal lymphoid tissues
- PMID: 10809967
- PMCID: PMC2326983
- DOI: 10.1046/j.1365-2567.2000.00013.x
Antibody repertoire development in fetal and newborn piglets, III. Colonization of the gastrointestinal tract selectively diversifies the preimmune repertoire in mucosal lymphoid tissues
Abstract
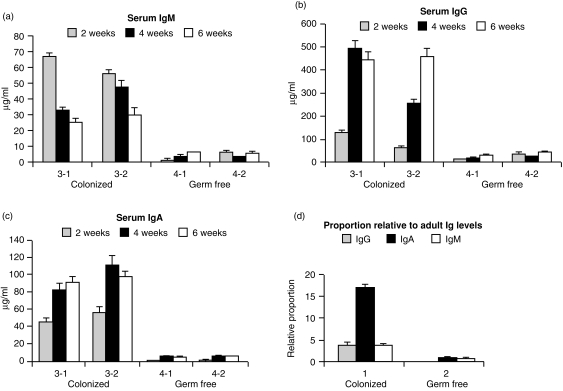
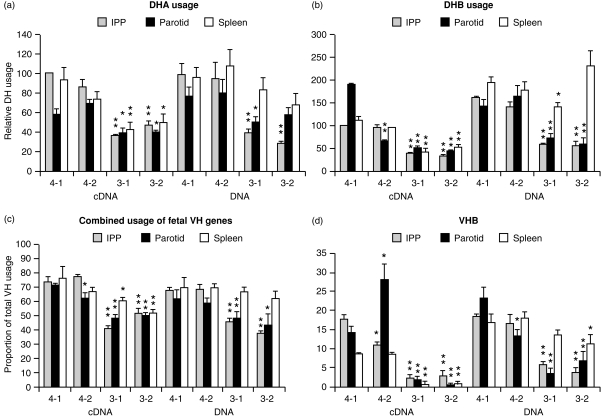
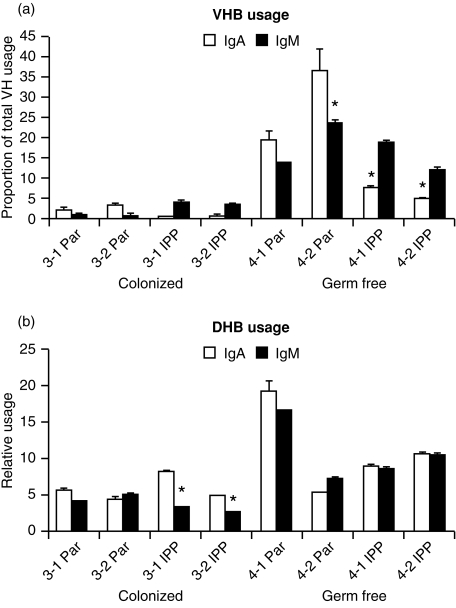
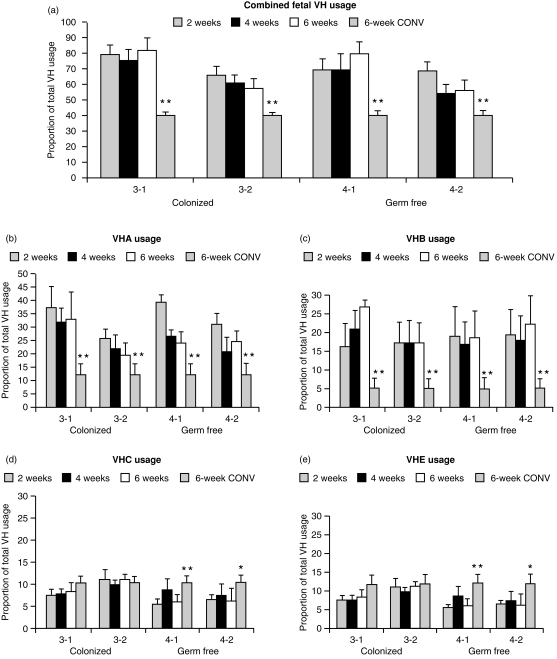
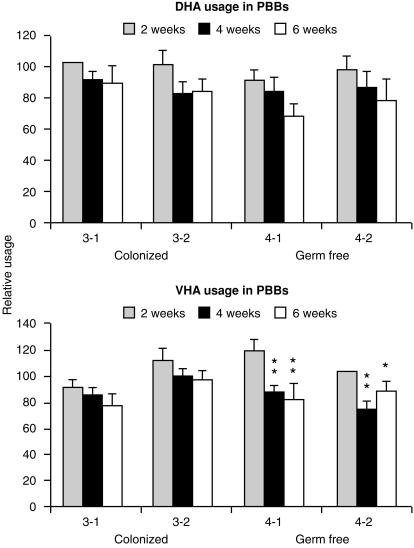
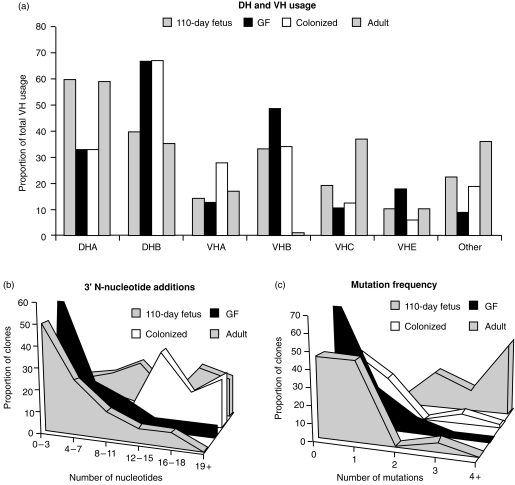
Changes in the VH-region repertoire of isolator piglets reared for 6 weeks under germ-free (GF) conditions and those colonized (COL) with a defined exclusion flora on the 1st day of life were compared. Although serum immunoglobulin levels were 20-100-fold higher in COL piglets than GF piglets, an analysis of peripheral blood B cells (PBBs) indicated that: GF and COL piglets used the same four VH genes and two DH segments during the 6-week period; proportional usage of VH genes and DH segments was the same as in fetal animals; and VH and DH usage did not differ between COL and GF animals. This pattern differed from the PBBs from 6-week-old conventional (CONV) piglets. When the sequences of 73 splenic CDR3 segments were analysed, DH usage and mutation frequency were the same in sequences from both 6-week-old GF and COL piglets; mutations were infrequent and occurred with the same frequency as in 110-day fetal spleen. However, the median CDR3 length in COL piglets was shifted upward due to 3' DH N-nucleotide additions. Neither COL nor GF animals made specific serum antibodies to phosphoryl choline given parenterally on a T-cell dependent carrier. In contrast to the near absence of a colonization effect in PBBs and splenic DNA, rearranged variable heavy-chain gene segments (VDJs) recovered from the DNA of mucosal lymphoid tissues of COL piglets showed pronounced differences from those recovered from GF animals in usage of DHA-, DHB-and VHB- and in the frequency of point mutation. The mucosal VDJ transcripts and those from the spleen were similarly affected by colonization. This effect on mucosal lymphoid tissue was consistent with the five-fold selective increase in serum immunoglobulin A (IgA) levels relative to IgM and IgG. Comparison of IgM and IgA transcripts from mucosal tissues suggested that IgA and IgM clones diversify in parallel. Our findings are the first to show that colonization of the gastrointestinal tract of offspring separated from their mothers, differs from 'conventionalized' GF animals in that colonization preferentially influences diversification and expansion of the preimmune IgM and IgA repertoire in mucosal lymphoid tissues but not in PBBs and seldom/modestly in VDJs from splenic DNA.
Figures

Similar articles
-
Antibody repertoire development in fetal and neonatal piglets. I. Four VH genes account for 80 percent of VH usage during 84 days of fetal life.J Immunol. 1998 Nov 1;161(9):5070-8. J Immunol. 1998. PMID: 9794445
-
Antibody repertoire development in fetal and neonatal piglets. XI. The thymic B-cell repertoire develops independently from that in blood and mesenteric lymph nodes.Immunology. 2005 Feb;114(2):171-83. doi: 10.1111/j.1365-2567.2004.02101.x. Immunology. 2005. PMID: 15667562 Free PMC article.
-
Antibody repertoire development in fetal and neonatal piglets. IV. Switch recombination, primarily in fetal thymus, occurs independent of environmental antigen and is only weakly associated with repertoire diversification.J Immunol. 2001 Sep 15;167(6):3239-49. doi: 10.4049/jimmunol.167.6.3239. J Immunol. 2001. PMID: 11544311
-
The VH and CH immunoglobulin genes of swine: implications for repertoire development.Vet Immunol Immunopathol. 1996 Nov;54(1-4):7-17. doi: 10.1016/s0165-2427(96)05680-2. Vet Immunol Immunopathol. 1996. PMID: 8988845 Review.
-
Antibody repertoire development in swine.Dev Comp Immunol. 2006;30(1-2):199-221. doi: 10.1016/j.dci.2005.06.025. Dev Comp Immunol. 2006. PMID: 16168480 Review.
Cited by
-
Neonatal colonisation expands a specific intestinal antigen-presenting cell subset prior to CD4 T-cell expansion, without altering T-cell repertoire.PLoS One. 2012;7(3):e33707. doi: 10.1371/journal.pone.0033707. Epub 2012 Mar 19. PLoS One. 2012. PMID: 22442714 Free PMC article.
-
Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain.J Virol. 2002 May;76(9):4241-50. doi: 10.1128/jvi.76.9.4241-4250.2002. J Virol. 2002. PMID: 11932389 Free PMC article.
-
Rearing environment affects development of the immune system in neonates.Clin Exp Immunol. 2010 Jun;160(3):431-9. doi: 10.1111/j.1365-2249.2010.04090.x. Epub 2010 Feb 22. Clin Exp Immunol. 2010. PMID: 20184618 Free PMC article.
-
Somatically diversified and proliferating transitional B cells: implications for peripheral B cell homeostasis.J Immunol. 2011 Jun 1;186(11):6437-44. doi: 10.4049/jimmunol.1003897. Epub 2011 Apr 27. J Immunol. 2011. PMID: 21525392 Free PMC article.
-
Generation of antibody- and B cell-deficient pigs by targeted disruption of the J-region gene segment of the heavy chain locus.Transgenic Res. 2011 Jun;20(3):625-41. doi: 10.1007/s11248-010-9444-z. Epub 2010 Sep 26. Transgenic Res. 2011. PMID: 20872248 Free PMC article.
References
-
- Kenworthy R, Crabb WE. Intestinal flora of young pigs, with reference to early weaning, Escherichia coli and scowers. J Comp Pathol. 1963;73:215. - PubMed
-
- Drasar BS, Barrow PA. Aspects of Microbiology 10: Intestinal Microbiology. Wokingham, UK: Van Nostrand Reinhold Ltd; 1985.
-
- Smith HW, Jones JET. The effect of the addition of copper sulfate to the diet on the bacterial flora of the alimentary tract of the pig. J Appl Bact. 1963;26:262.
-
- Tannock GW. The normal microflora: new concepts in health promotion. Microbiol Sci. 1988;5:4. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
