

Structural and functional conservation at the boundaries of the chicken beta-globin domain
- PMID: 10811622
- PMCID: PMC384375
- DOI: 10.1093/emboj/19.10.2315
Structural and functional conservation at the boundaries of the chicken beta-globin domain
Abstract
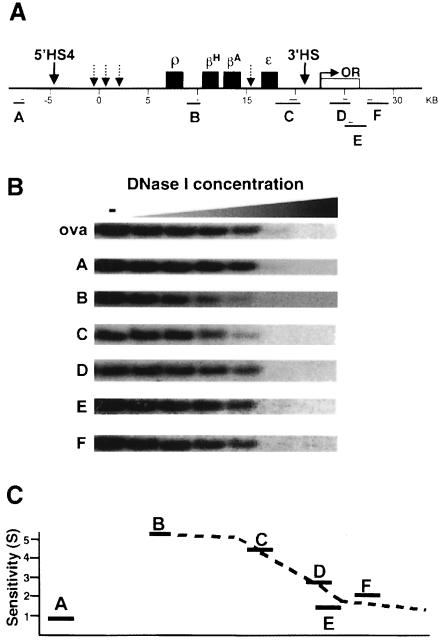
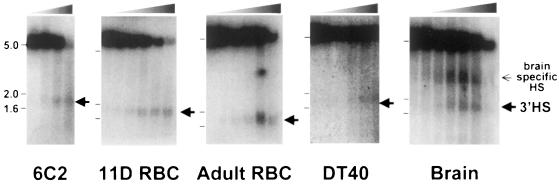
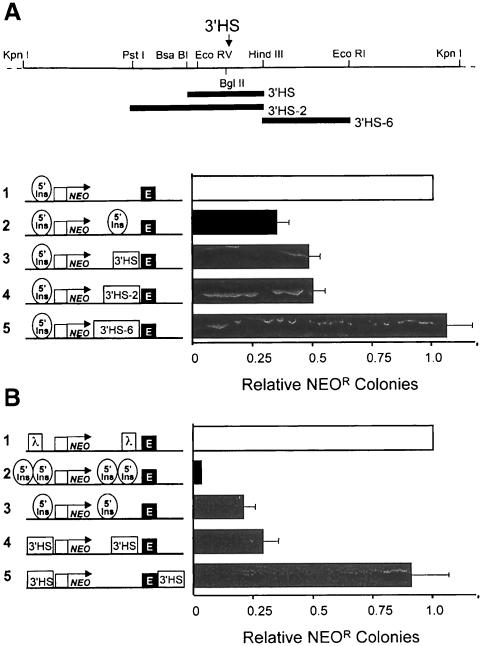
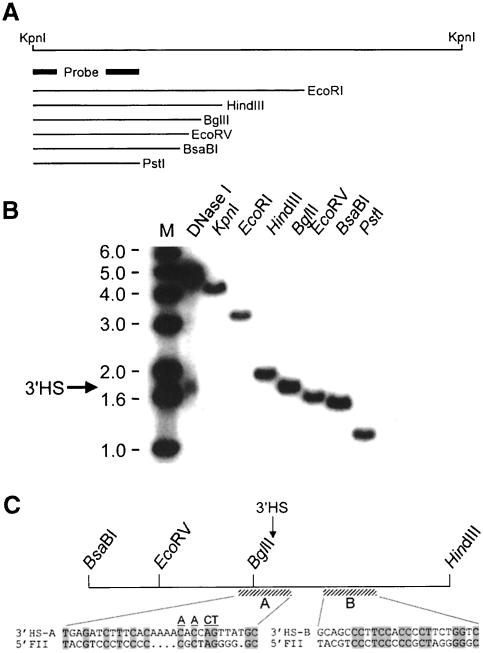
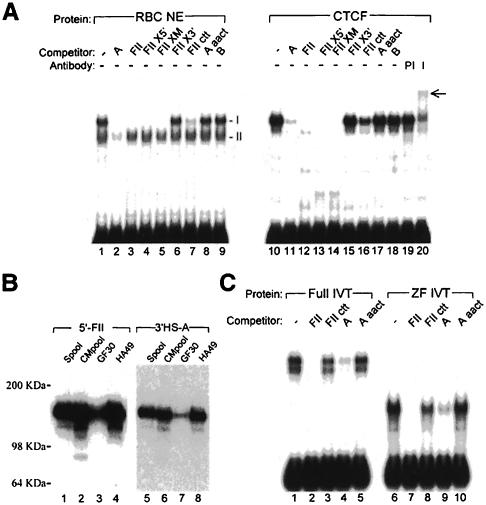
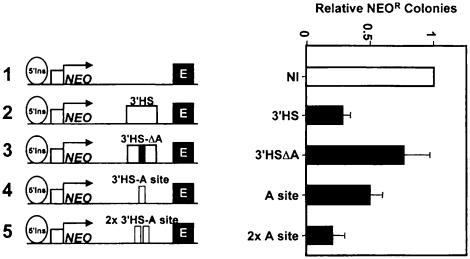
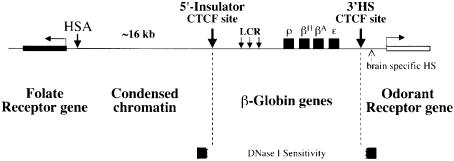
We show that the 3' boundary of the chicken beta-globin locus bears striking structural similarities to the 5' boundary. In erythroid cells a clear transition in DNase I sensitivity of chromatin at the 3' end of the locus is observed, the location of this transition is marked by a constitutive DNase I hypersensitive site (HS), and DNA spanning this site has the enhancer-blocking capacity of an insulator. This HS contains a binding site for the transcription factor CTCF. As in the case of the 5' insulator, the CTCF site is both necessary and sufficient for the enhancer-blocking activity of the 3' boundary. The position of this insulator is consistent with our proposal that it may function to maintain the distinct regulatory programs of the globin genes and their closely appended 3' neighbor, an odorant receptor gene. We conclude that both boundaries of the chicken beta-globin domain are capable of playing functionally similar roles and that the same protein is a necessary component of the molecular mechanism through which these boundaries are defined.
Figures
References
-
- Abe K., Kusakabe,Y., Tanemura,K., Emori,Y. and Arai,S. (1993) Multiple genes for G protein-coupled receptors and their expression in lingual epithelia. FEBS Lett., 316, 253–256. - PubMed
-
- Bell A.C. and Felsenfeld,G. (1999) Stopped at the border: boundaries and insulators. Curr. Opin. Genet. Dev., 9, 191–198. - PubMed
-
- Bell A.C., West,A.G. and Felsenfeld,G. (1999) The protein CTCF is required for the enhancer-blocking activity of vertebrate insulators. Cell, 98, 387–396. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
