

The capsule is a virulence determinant in the pathogenesis of Pasteurella multocida M1404 (B:2)
- PMID: 10816499
- PMCID: PMC97626
- DOI: 10.1128/IAI.68.6.3463-3468.2000
The capsule is a virulence determinant in the pathogenesis of Pasteurella multocida M1404 (B:2)
Abstract
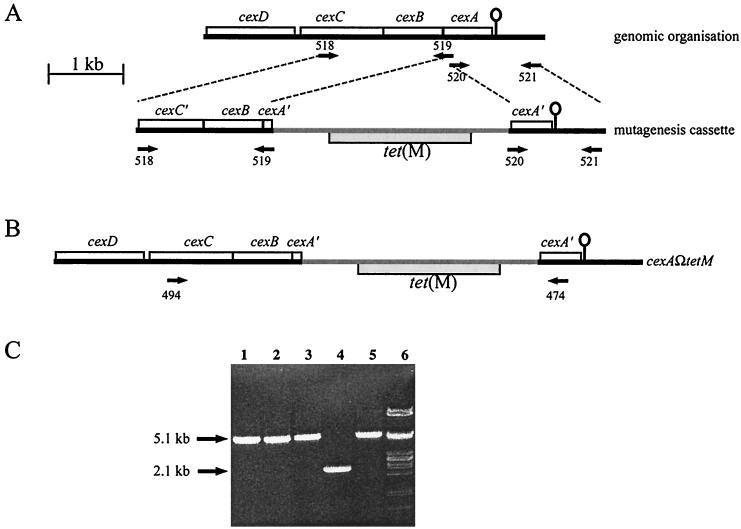
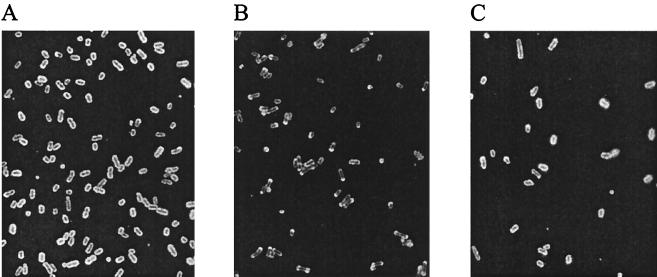
Capsules from a range of pathogenic bacteria are key virulence determinants, and the capsule has been implicated in virulence in Pasteurella multocida. We have previously identified and determined the nucleotide sequence of the P. multocida M1404 (B:2) capsule biosynthetic locus (J. D. Boyce, J. Y. Chung, and B. Adler, Vet. Microbiol. 72:121-134, 2000). The cap locus consists of 15 genes, which can be grouped into three functional regions. Regions 1 and 3 contain genes proposed to encode proteins involved in capsule export, and region 2 contains genes proposed to encode proteins involved in polysaccharide biosynthesis. In order to construct a mutant impaired in capsule export, the final gene of region 1, cexA, was disrupted by insertion of a tetracycline resistance cassette by allelic replacement. The genotype of the tet(M) OmegacexA mutant was confirmed by Southern hybridization and PCR. The acapsular phenotype was confirmed by immunofluorescence, and the strain could be complemented and returned to capsule production by the presence of a cloned uninterrupted copy of cexA. Wild-type, mutant, and complemented strains were tested for virulence by intraperitoneal challenge of mice; the presence of the capsule was shown to be a crucial virulence determinant. Following intraperitoneal challenge of mice, the acapsular bacteria were removed efficiently from the blood, spleen, and liver, while wild-type bacteria multiplied rapidly. Acapsular bacteria were readily taken up by murine peritoneal macrophages, but wild-type bacteria were significantly resistant to phagocytosis. Both wild-type and acapsular bacteria were resistant to complement in bovine and murine serum.
Figures
References
-
- Anderson L C, Rush H G, Glorioso J C. Strain differences in the susceptibility and resistance of Pasteurella multocida to phagocytosis and killing by rabbit polymorphonuclear neutrophils. Am J Vet Res. 1984;45:1193–1198. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: Greene Publishing Associates and Wiley-Interscience; 1987.
-
- Boyce J D, Chung J Y, Adler B. Genetic organisation of the capsule biosynthetic locus of Pasteurella multocida M1404 (B:2) Vet Microbiol. 2000;72:121–134. - PubMed
-
- Carter G R. Pasteurellosis: Pasteurella multocida and Pasteurella hemolytica. Adv Vet Sci. 1967;11:321–379. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
