

The something about silencing protein, Sas3, is the catalytic subunit of NuA3, a yTAF(II)30-containing HAT complex that interacts with the Spt16 subunit of the yeast CP (Cdc68/Pob3)-FACT complex
- PMID: 10817755
- PMCID: PMC316621
The something about silencing protein, Sas3, is the catalytic subunit of NuA3, a yTAF(II)30-containing HAT complex that interacts with the Spt16 subunit of the yeast CP (Cdc68/Pob3)-FACT complex
Abstract
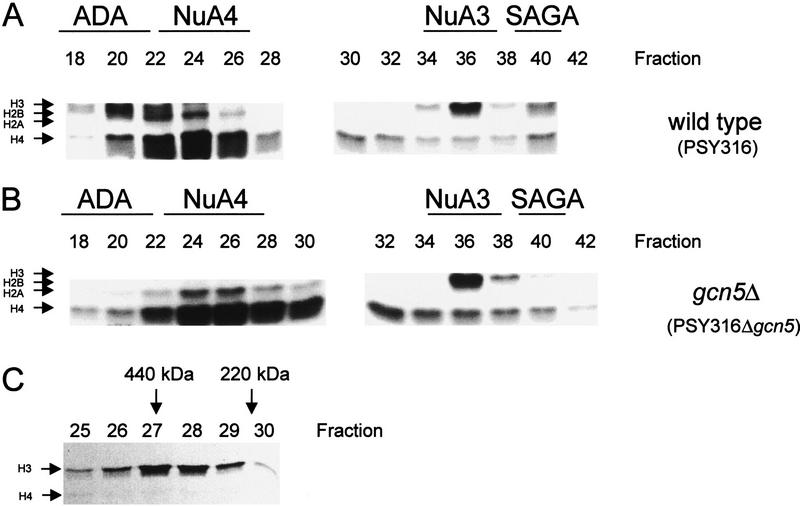
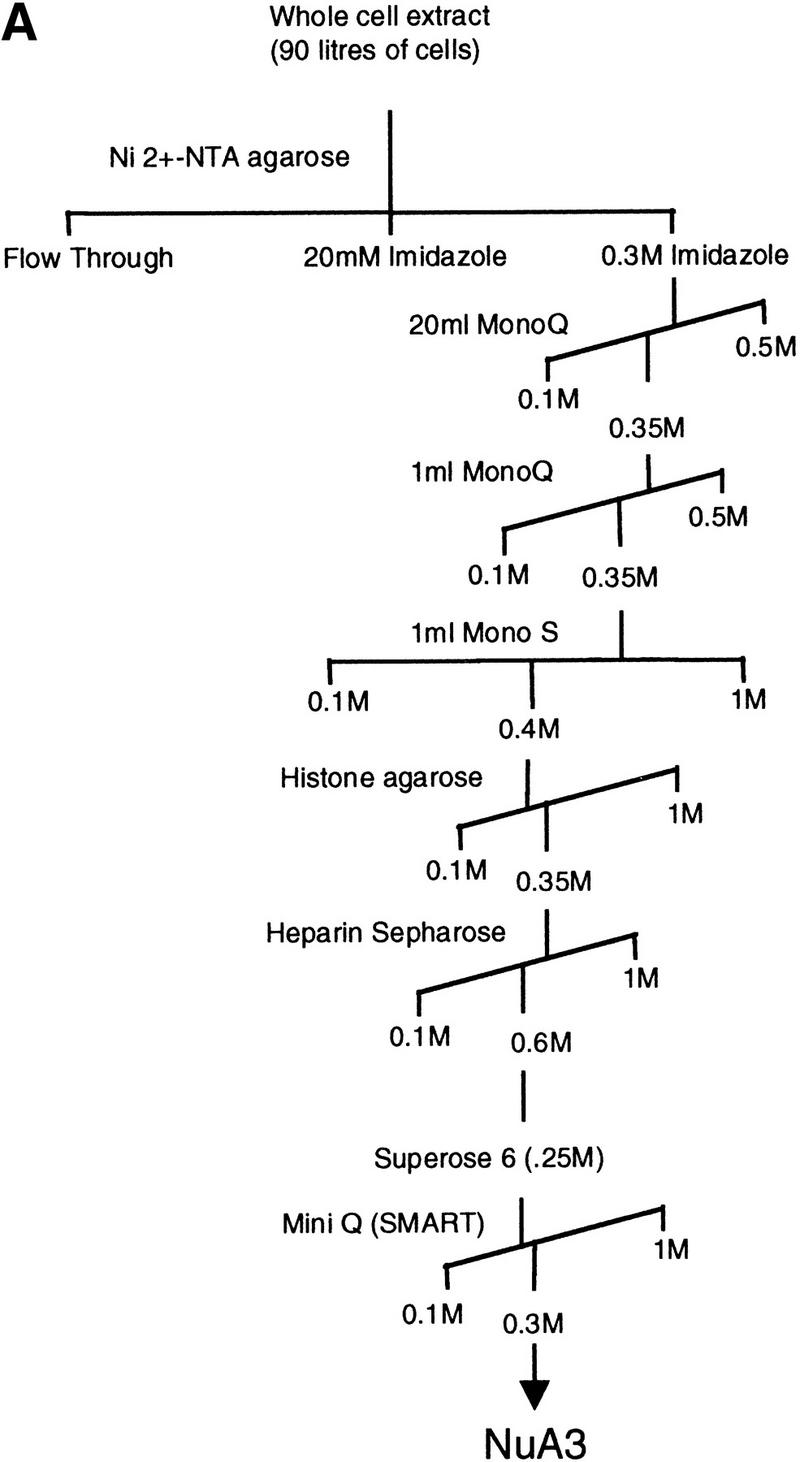
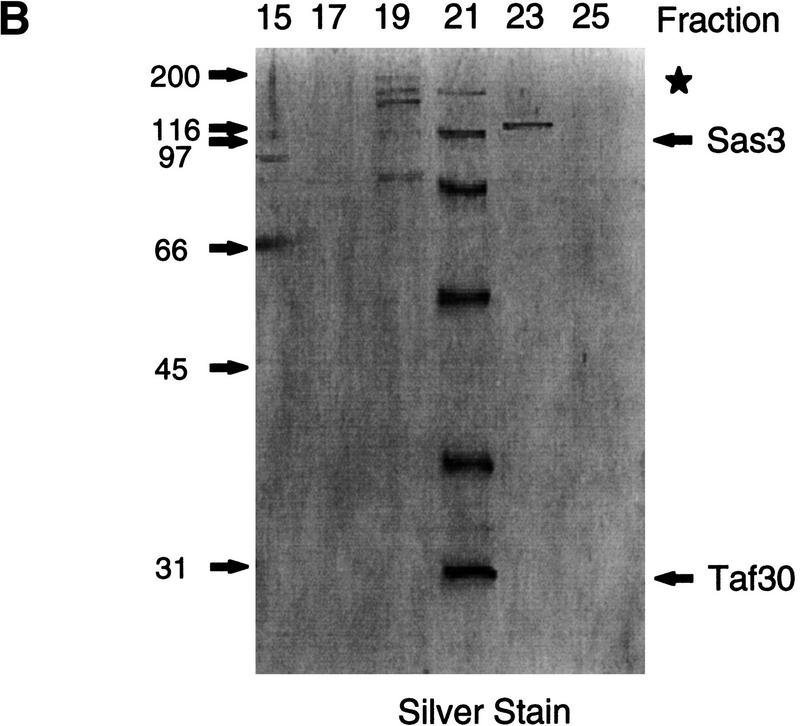
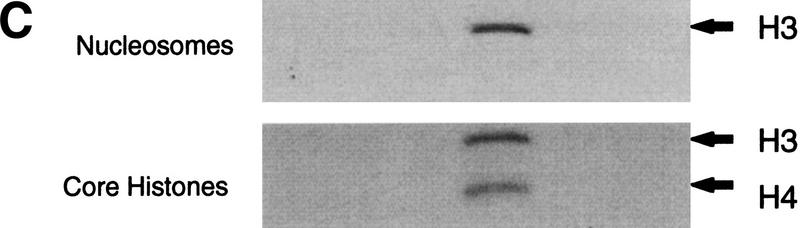
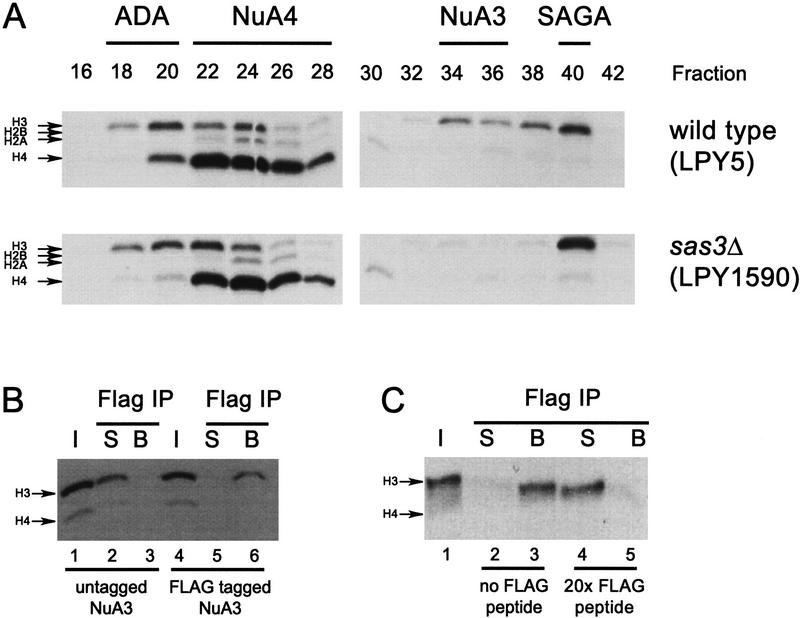
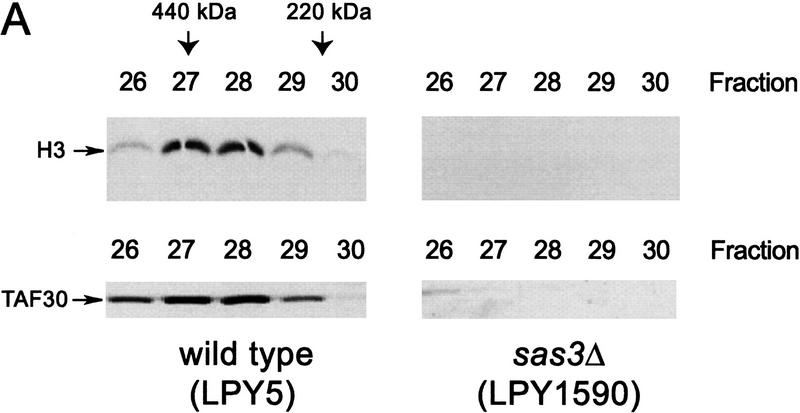
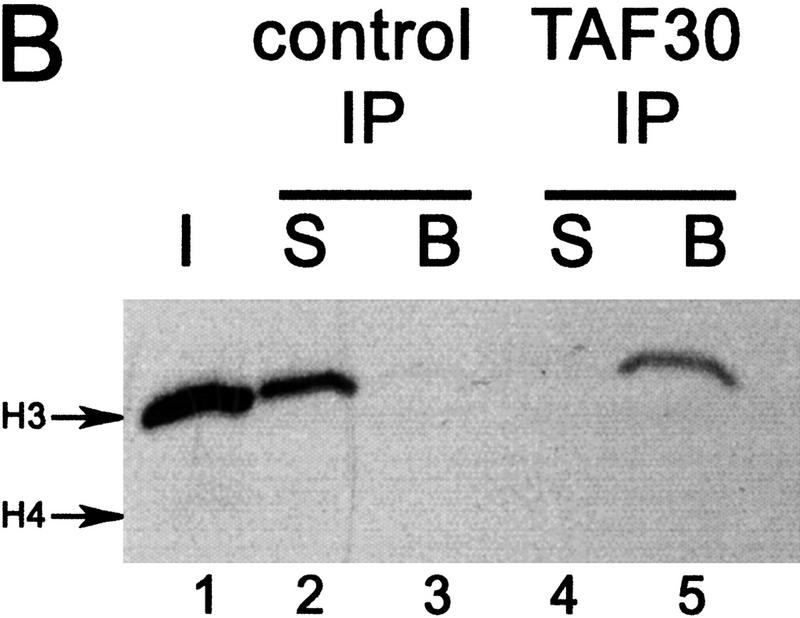
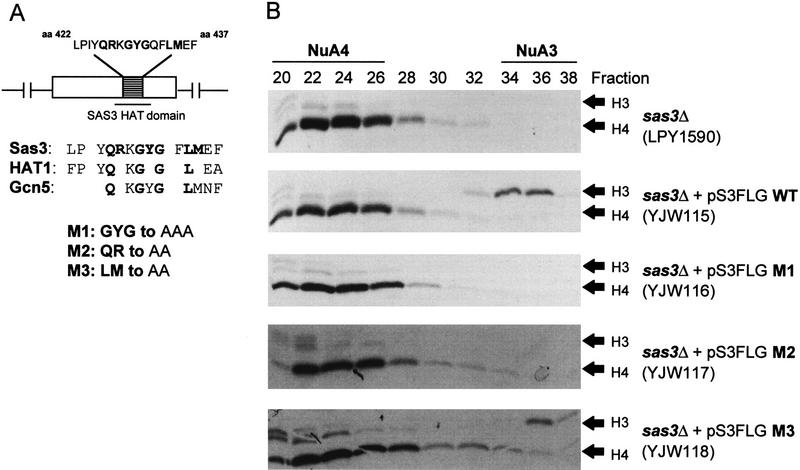
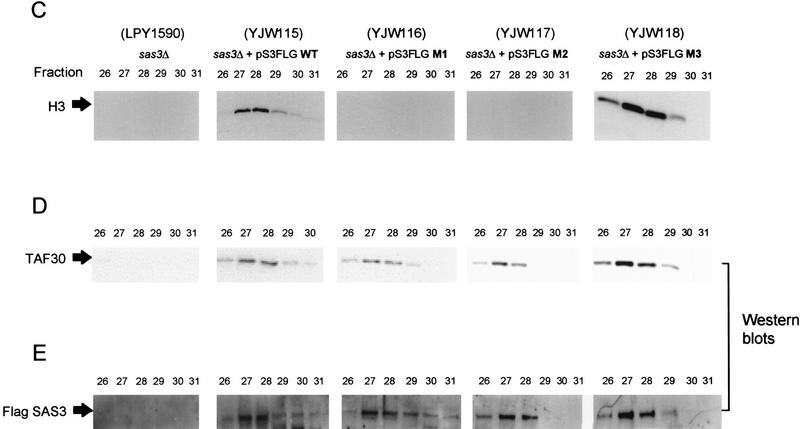
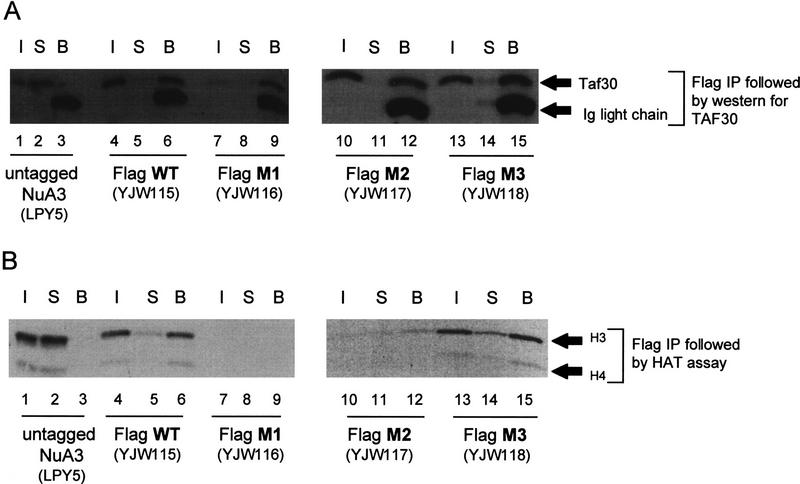
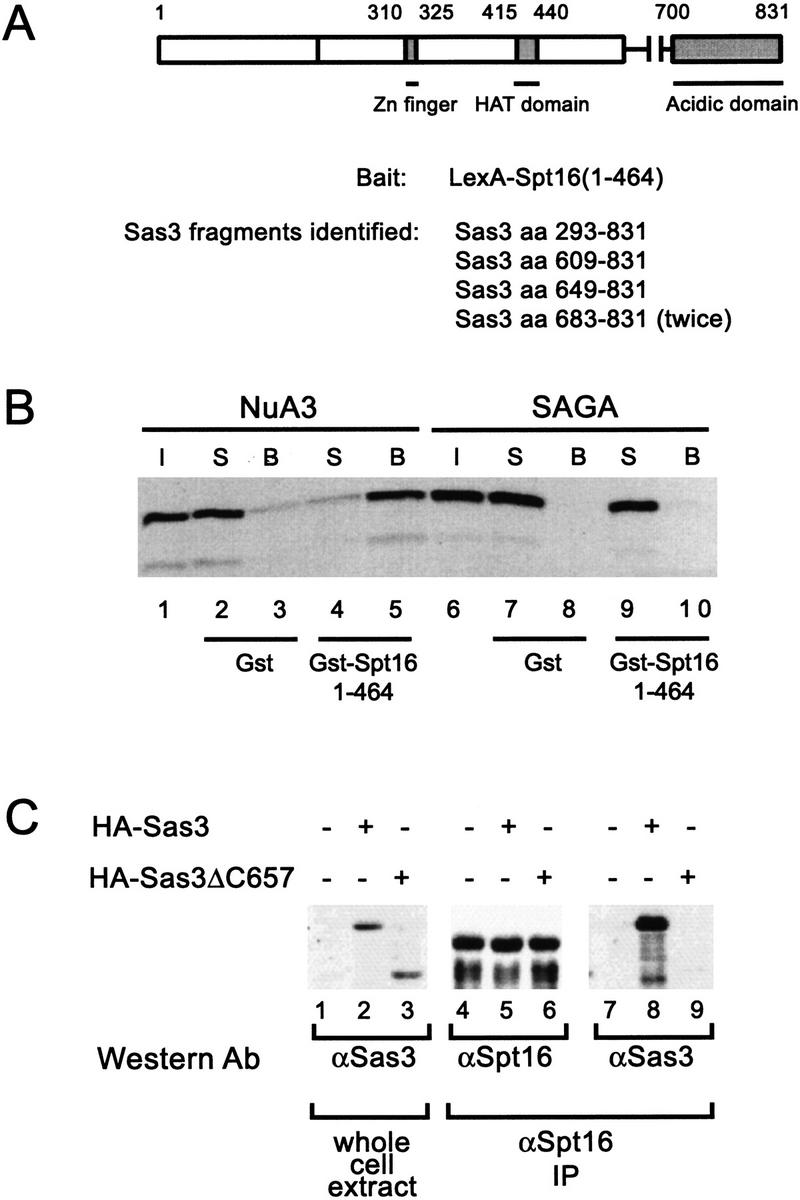
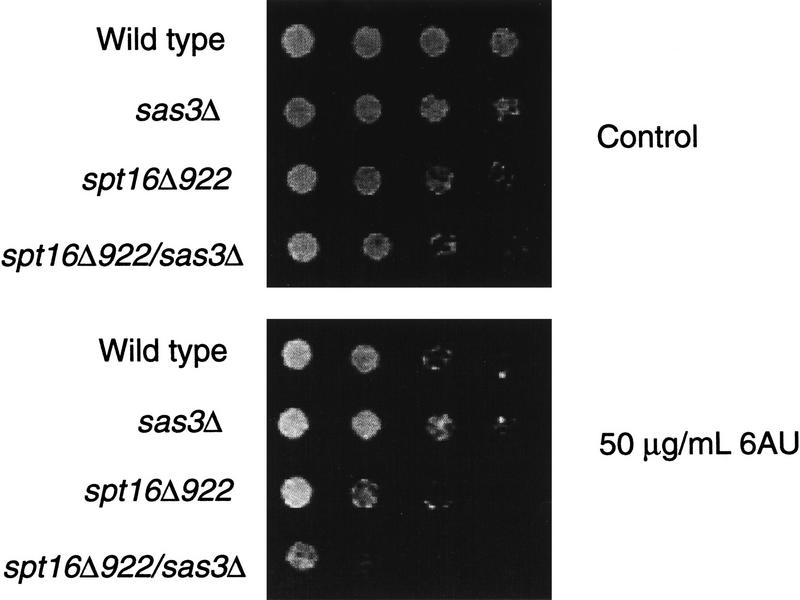
We have purified and characterized a Gcn5-independent nucleosomal histone H3 HAT complex, NuA3 (Nucleosomal Acetyltransferase of histone H3). Peptide sequencing of proteins from the purified NuA3 complex identified Sas3 as the catalytic HAT subunit of the complex. Sas3 is the yeast homolog of the human MOZ oncogene. Sas3 is required for both the HAT activity and the integrity of the NuA3 complex. In addition, NuA3 contains the TBP- associated factor, yTAF(II)30, which is also a component of the TFIID, TFIIF, and SWI/SNF complexes. Sas3 mediates interaction of the NuA3 complex with Spt16 both in vivo and in vitro. Spt16 functions as a component of the yeast CP (Cdc68/Pob3) and mammalian FACT (facilitates chromatin transcription) complexes, which are involved in transcription elongation and DNA replication. This interaction suggests that the NuA3 complex might function in concert with FACT-CP to stimulate transcription or replication elongation through nucleosomes by providing a coupled acetyltransferase activity.
Figures
References
-
- Ausubel FM. Short protocols in molecular biology: A compendium of methods from Current protocols in molecular biology. NY: John Wiley & Sons; 1995.
-
- Bannister AJ, Kouzarides T. The CBP coactivator is a histone acetyltransferase. Nature. 1996;384:641–643. - PubMed
-
- Borrow J, Stanton VPJ, Andresen JM, Becher RB, Chaganti FG, RS, Civin CI, Disteche C, Dube I, Frischauf AM, Horsman D, Mitelman F, Volinia S, Watmore AE, Housman DE. The translocation t(8;16)(p11;p13) of acute myeloid leukaemia fuses a putative acetyltransferase to the CREB-binding protein. Nat Genet. 1996;14:33–41. - PubMed
-
- Brewster NK, Johnston GC, Singer RA. Characterization of the CP complex, an abundant dimer of Cdc68 and Pob3 proteins that regulates yeast transcriptional activation and chromatin repression. J Biol Chem. 1998;273:21972–21979. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous