

Intracellular modulation of NMDA receptor function by antipsychotic drugs
- PMID: 10818136
- PMCID: PMC4203343
- DOI: 10.1523/JNEUROSCI.20-11-04011.2000
Intracellular modulation of NMDA receptor function by antipsychotic drugs
Abstract
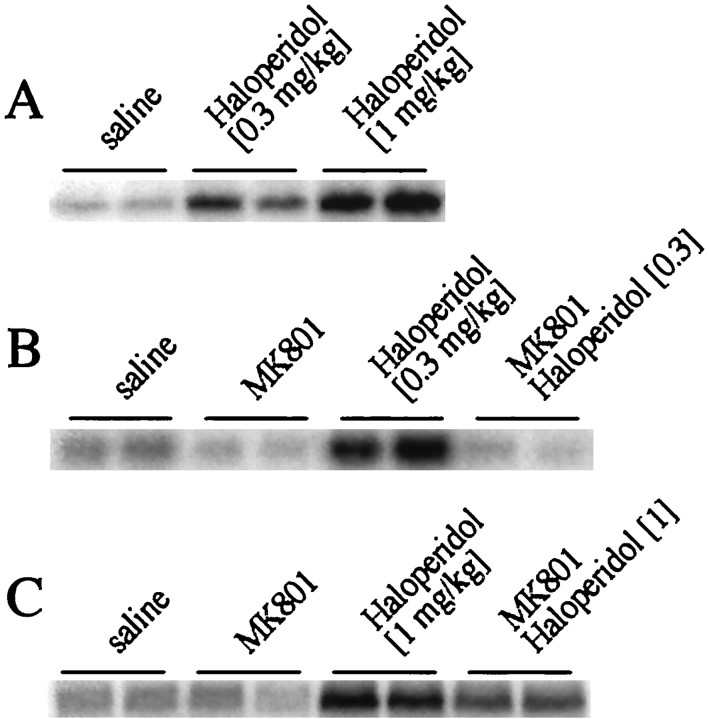
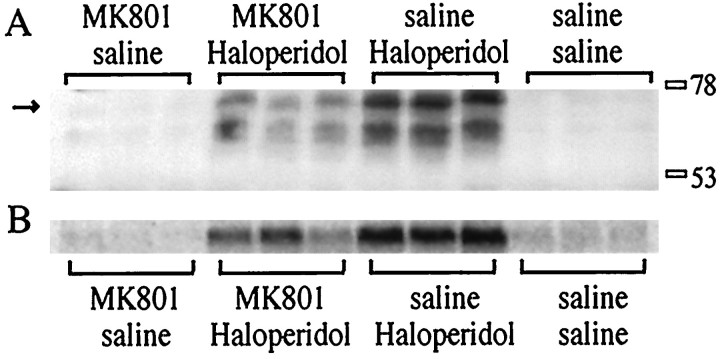
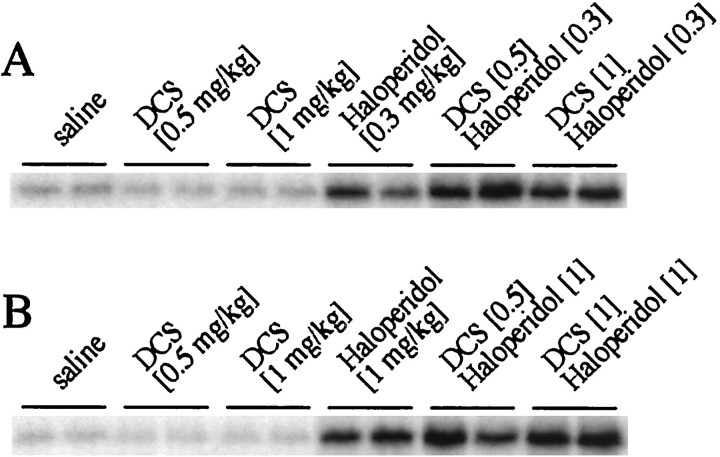
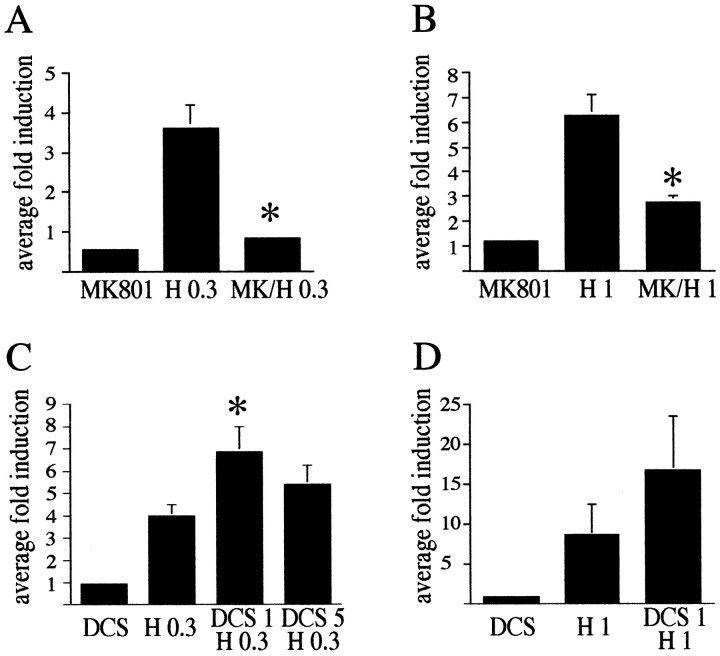
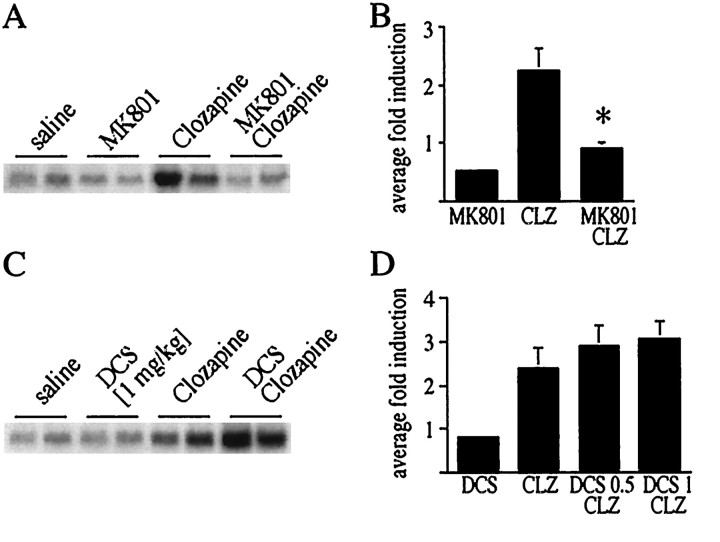
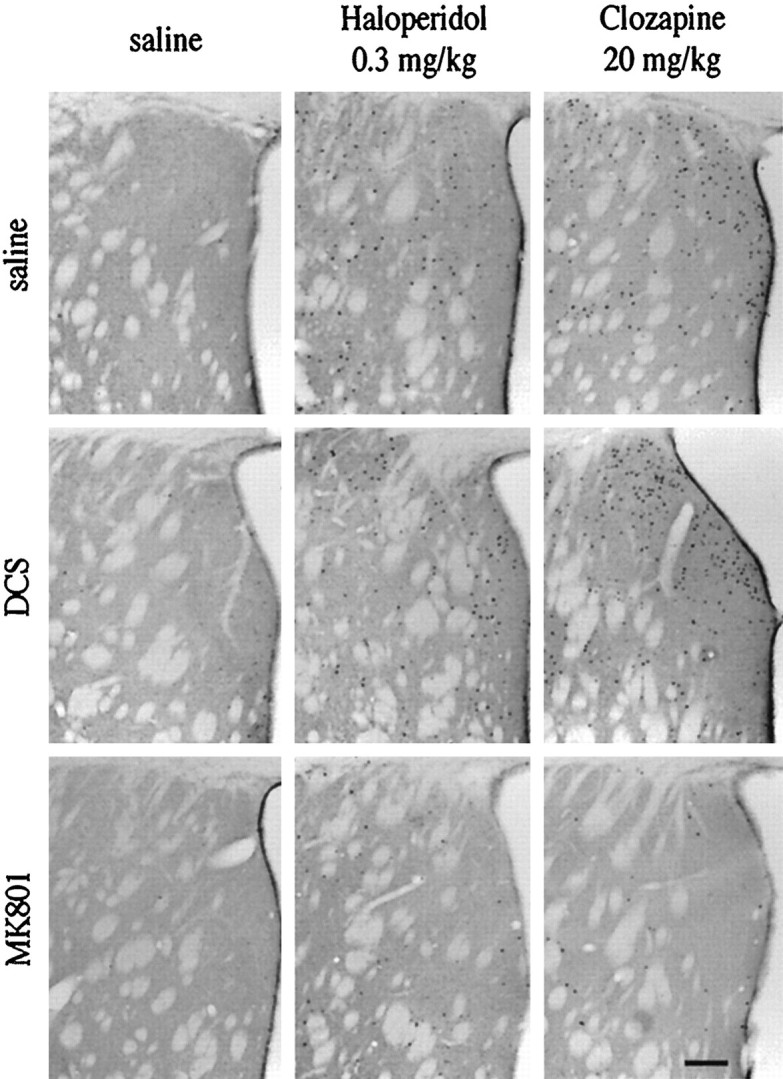
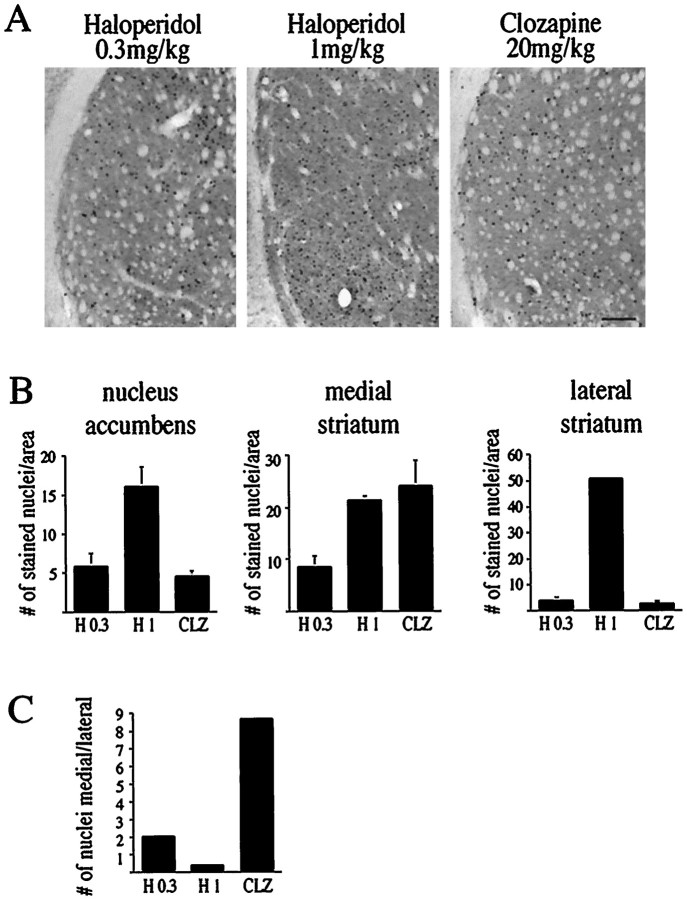
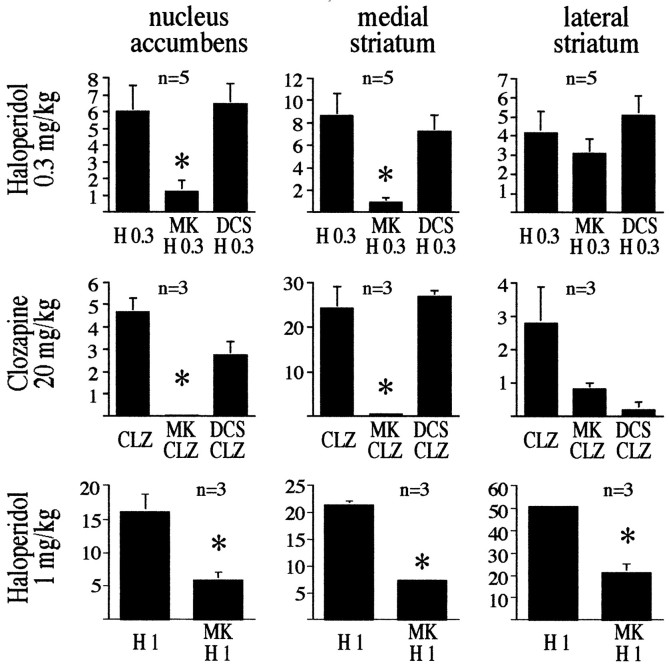
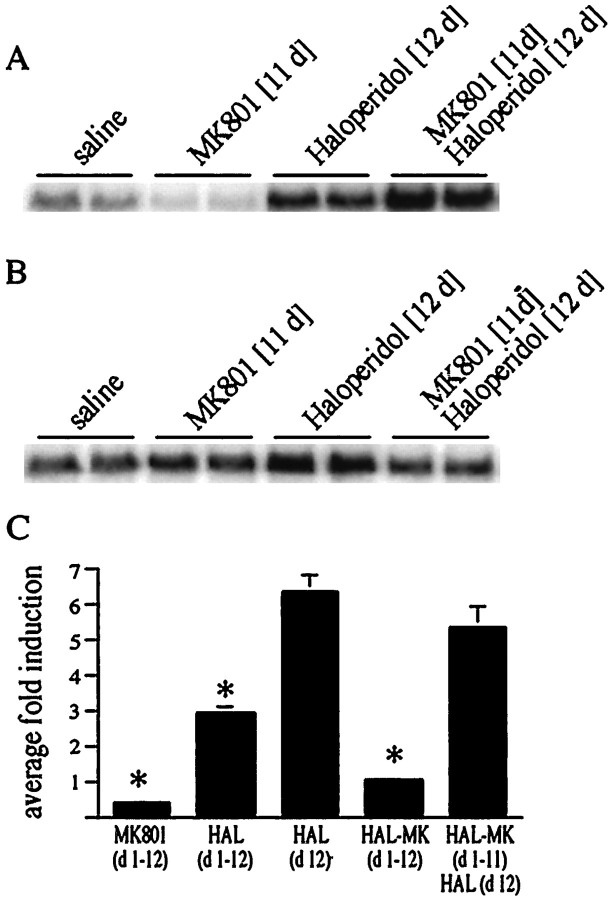
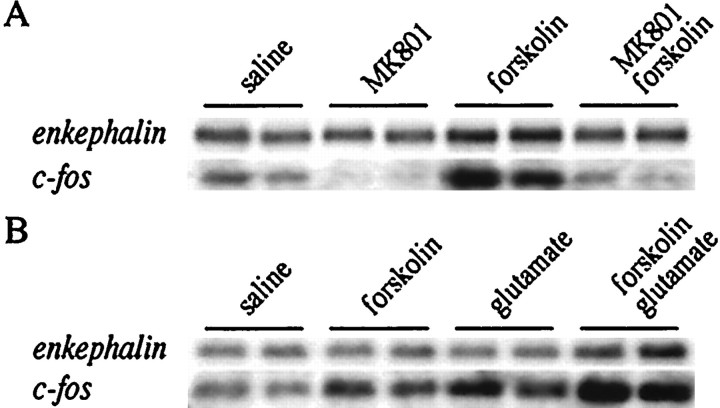
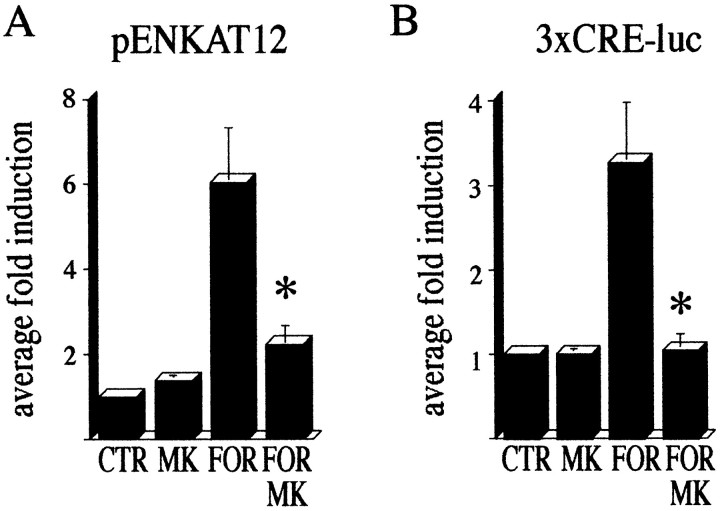
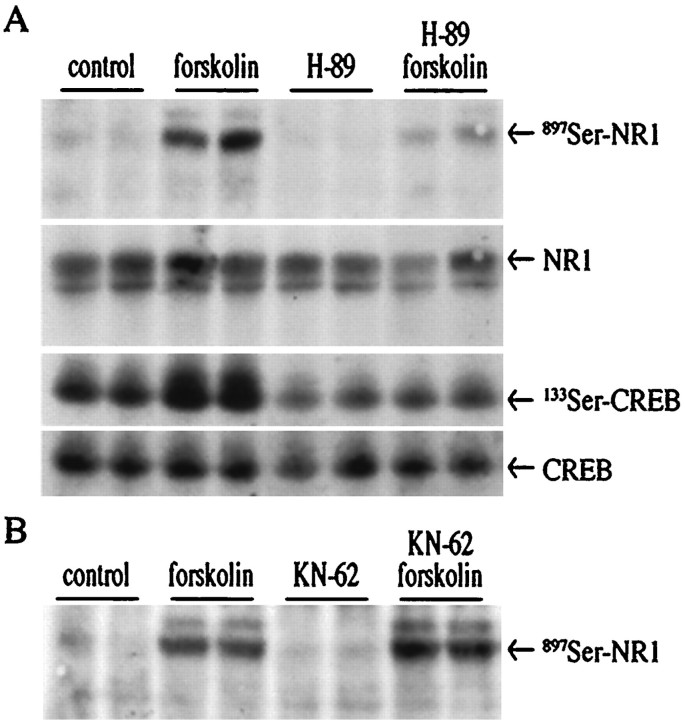
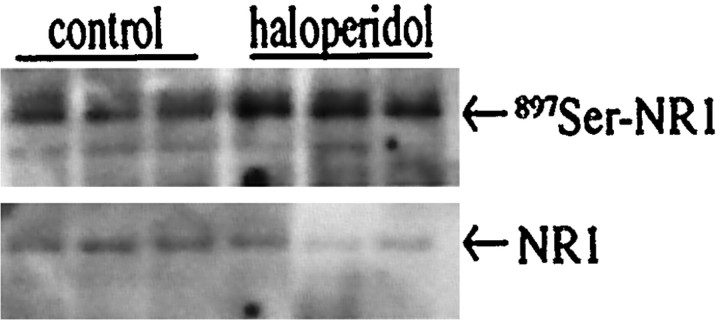
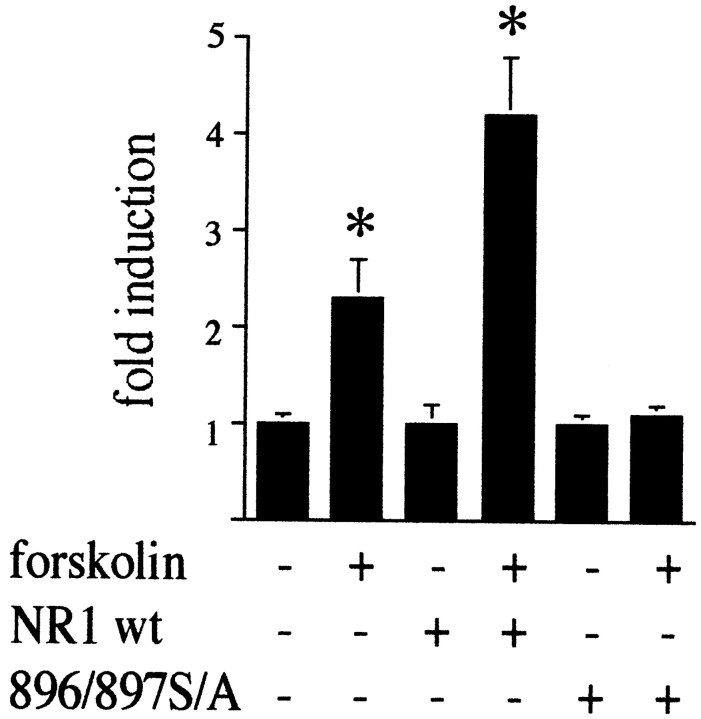
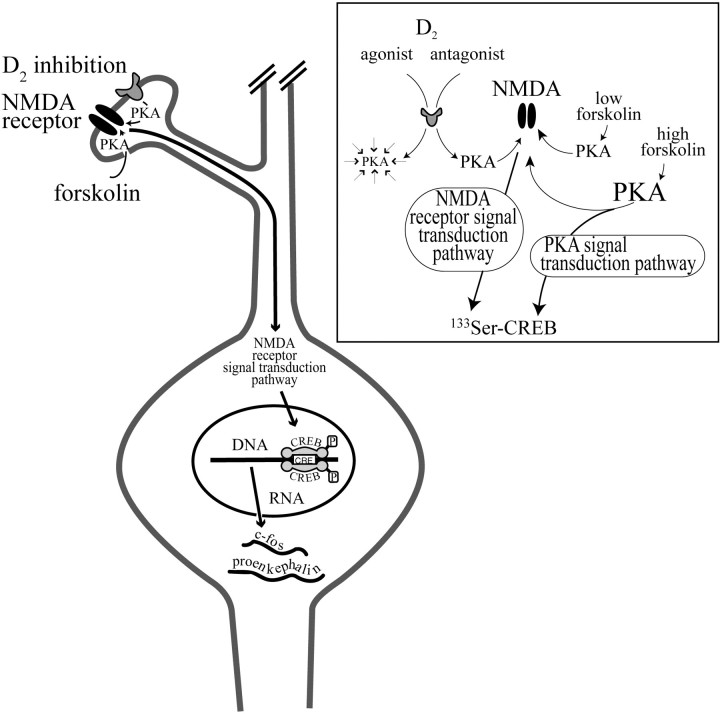
The present study deals with the functional interaction of antipsychotic drugs and NMDA receptors. We show that both the conventional antipsychotic drug haloperidol and the atypical antipsychotic drug clozapine mediate gene expression via intracellular regulation of NMDA receptors, albeit to different extents. Data obtained in primary striatal culture demonstrate that the intraneuronal signal transduction pathway activated by haloperidol, the cAMP pathway, leads to phosphorylation of the NR1 subtype of the NMDA receptor at (897)Ser. Haloperidol treatment is likewise shown to increase (897)Ser-NR1 phosphorylation in rats in vivo. Mutation of (896)Ser and (897)Ser to alanine, which prevents phosphorylation at both sites, inhibits cAMP-mediated gene expression. We conclude that antipsychotic drugs have the ability to modulate NMDA receptor function by an intraneuronal signal transduction mechanism. This facilitation of NMDA activity is necessary for antipsychotic drug-mediated gene expression and may contribute to the therapeutic benefits as well as side effects of antipsychotic drug treatment.
Figures
References
-
- Auchus AP, Pickel VM. Quantitative light microscopic demonstration of increased pallidal and striatal met5-enkephalin-like immunoreactivity in rats following chronic treatment with haloperidol but not with clozapine: implications for the pathogenesis of neuroleptic-induced movement disorders. Exp Neurol. 1992;117:17–27. - PubMed
-
- Banerjee SP, Zuck LG, Yablonsky-Alter E, Lidsky TI. Glutamate agonist activity: implications for antipsychotic drug action and schizophrenia. NeuroReport. 1995;6:2500–2504. - PubMed
-
- Berger SL, Chirgwin JM. Isolation of RNA. Methods Enzymol. 1989;180:3–13. - PubMed
-
- Boegman RJ, Vincent SR. Involvement of adenosine and glutamate receptors in the induction of c-fos in the striatum by haloperidol. Synapse. 1996;22:70–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources