

Exploration of the ligand binding site of the human 5-HT(4) receptor by site-directed mutagenesis and molecular modeling
- PMID: 10821780
- PMCID: PMC1572113
- DOI: 10.1038/sj.bjp.0703356
Exploration of the ligand binding site of the human 5-HT(4) receptor by site-directed mutagenesis and molecular modeling
Abstract
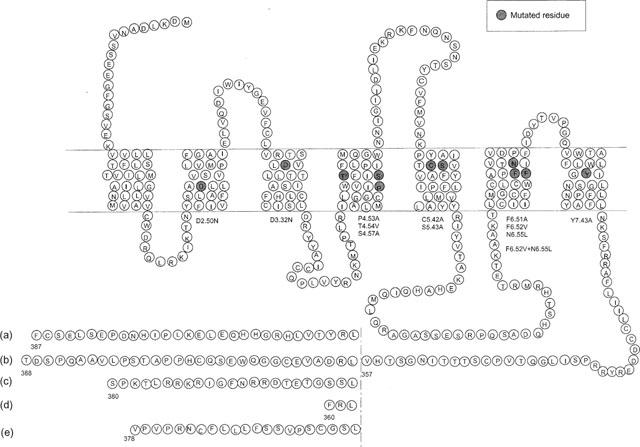
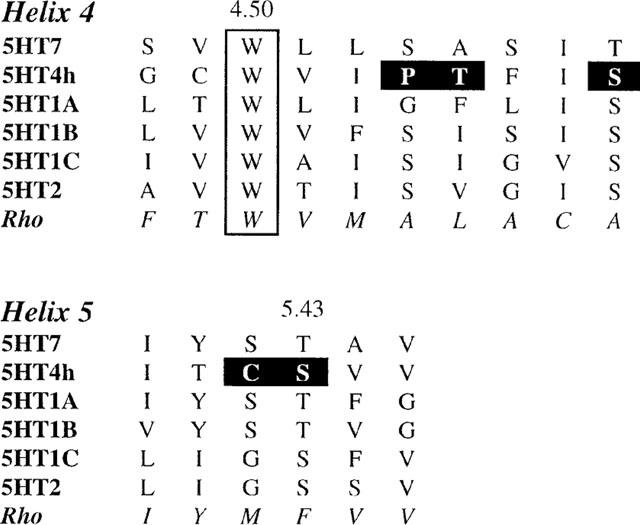
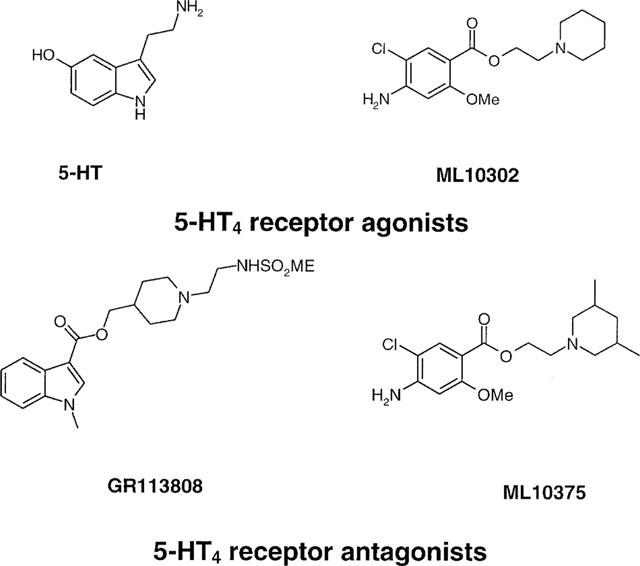
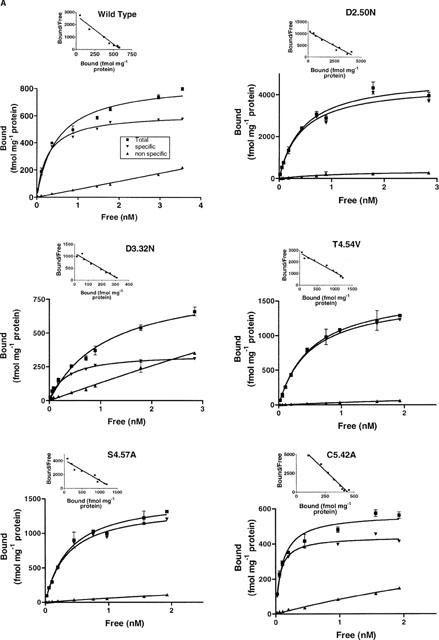
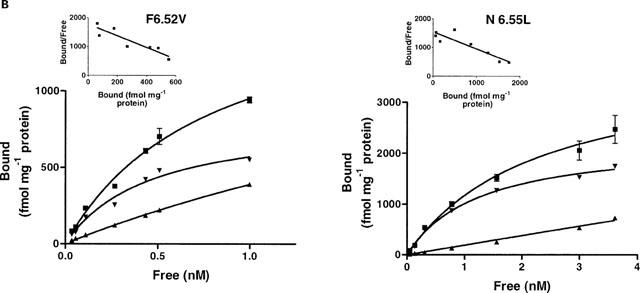
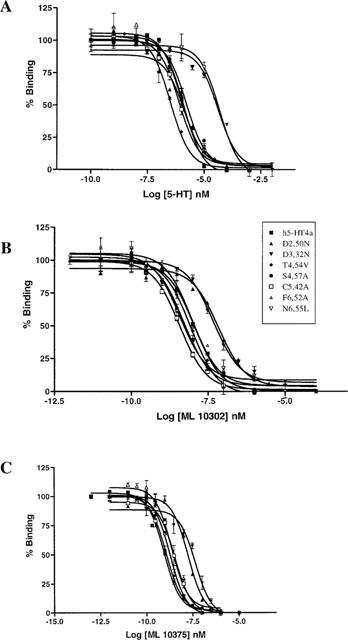
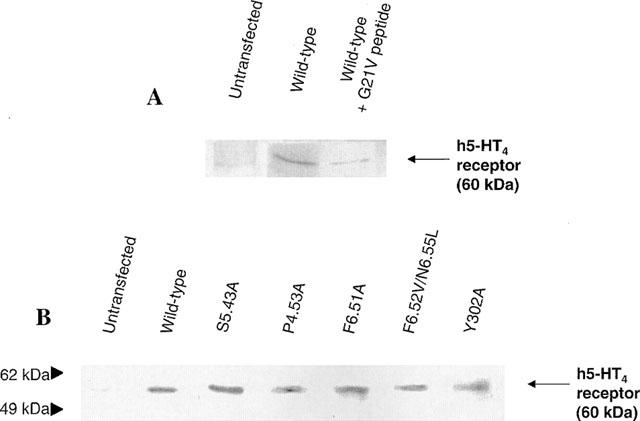
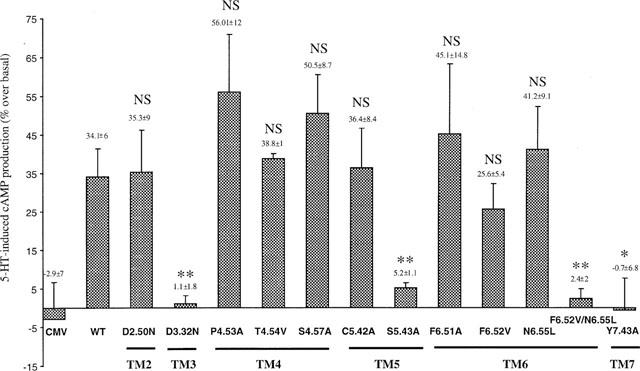
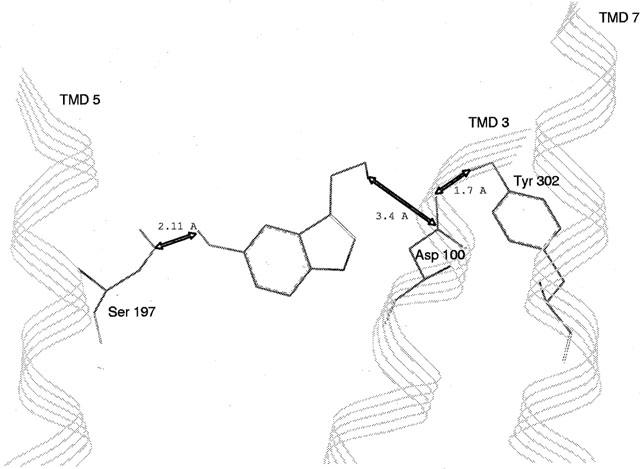
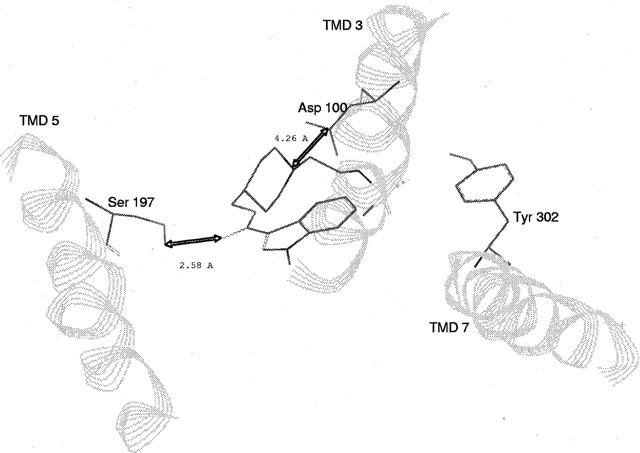
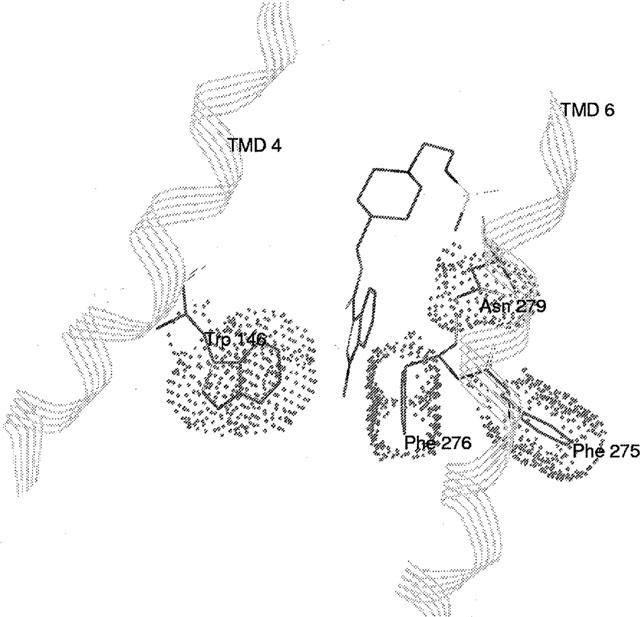
Among the five human 5-HT(4) (h5-HT(4)) receptor isoforms, the h5-HT(4(a)) receptor was studied with a particular emphasis on the molecular interactions involved in ligand binding. For this purpose, we used site-directed mutagenesis of the transmembrane domain. Twelve mutants were constructed with a special focus on the residue P4.53 of helix IV which substitutes in h5-HT(4) receptors the highly conserved S residue among the rhodopsin family receptors. The mutated receptors were transiently expressed in COS-7 cells. Ligand binding or competition studies with two h5-HT(4) receptor agonists, serotonin and ML10302 and two h5-HT(4) receptor antagonists, [(3)H]-GR113808 and ML10375 were performed on wild type and mutant receptors. Functional activity of the receptors was evaluated by measuring the ability of serotonin to stimulate adenylyl cyclase. Ligand binding experiments revealed that [(3)H]-GR113808 did not bind to mutants P4.53A, S5.43A, F6.51A, Y7.43A and to double mutant F6.52V/N6.55L. On the other hand mutations D3.32N, S5.43A and Y7.43A appeared to promote a dramatic decrease of h5-HT(4(a)) receptor functional activity. From these studies, S5.43 and Y7.43 clearly emerged as common anchoring sites to antagonist [(3)H]-GR113808 and to serotonin. According to these results, we propose ligand-receptor complex models with serotonin and [(3)H]-GR113808. For serotonin, three interaction points were selected including ionic interaction with D3.32, a stabilizing interaction of this ion pair by Y7.43 and a hydrogen bond with S5.43. [(3)H]-GR113808 was also docked, based on the same type of interactions with S5.43 and D3.32: the proposed model suggested a possible role of P4.53 in helix IV structure allowing the involvement of a close hydrophobic residue, W4.50, in a hydrophobic pocket for hydrophobic interactions with the indole ring of [(3)H]-GR113808.
Figures
References
-
- ALMAULA N., EBERSOLE B.J., BALLESTEROS J.A., WEINSTEIN H., SEALFON S.C. Contribution of a helix 5 locus selectivity of hallucinogenic and non-hallucinogenic ligands for the human 5-Hydroxytryptamine2A and 5-Hydroxytryptamine2C receptors: direct and indirect effects on ligand affinity mediated by the same locus. Mol. Pharmacol. 1996;50:34–42. - PubMed
-
- BALDWIN J.M. Structure and function of receptors coupled to G proteins. Curr. Opin. Cell. Biol. 1994;6:180–190. - PubMed
-
- BALDWIN J.M., SCHERTLER G.F.X., UNGER V.M. An alpha-carbon template for the transmembrane helices in rhodopsin family of G-protein-coupled receptors. J. Mol. Biol. 1997;272:144–164. - PubMed
-
- BALLESTEROS J.A., WEINSTEIN H. Integrated methods for modelling G-protein coupled receptors. Methods Neurosci. 1995;25:366–428.
-
- BLONDEL O., GASTINEAU M., DAHMOUNE Y., LANGLOIS M., FISCHMEISTER R. Cloning, expression, and pharmacology of four human 5-hydroxytryptamine4 receptor isoforms produced by alternative splicing in the carboxyl terminus. J. Neurochem. 1998;70:2252–2261. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
