

A mechanistic basis for Mre11-directed DNA joining at microhomologies
- PMID: 10823903
- PMCID: PMC18616
- DOI: 10.1073/pnas.110144297
A mechanistic basis for Mre11-directed DNA joining at microhomologies
Abstract
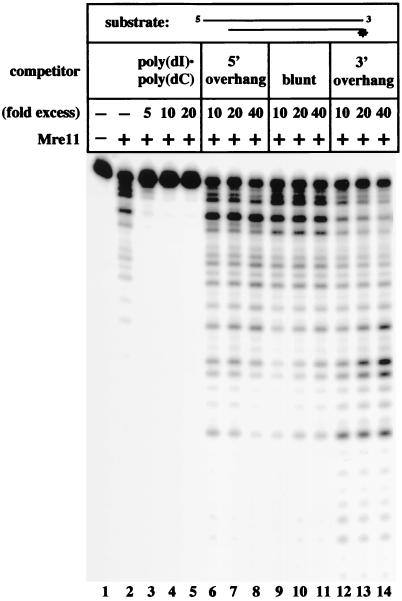
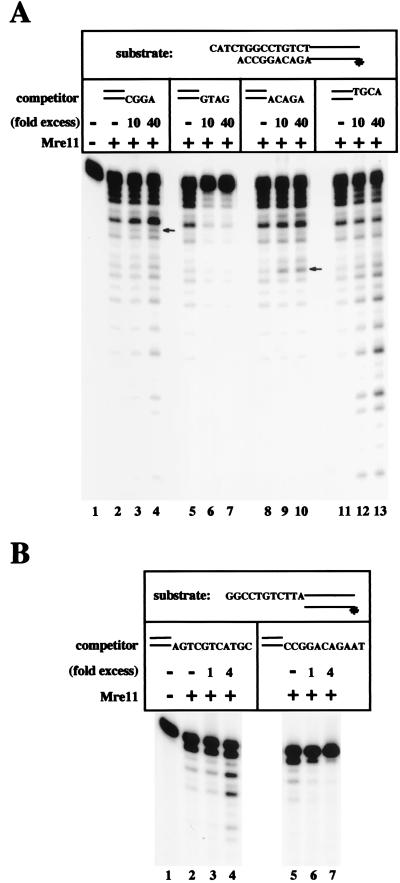
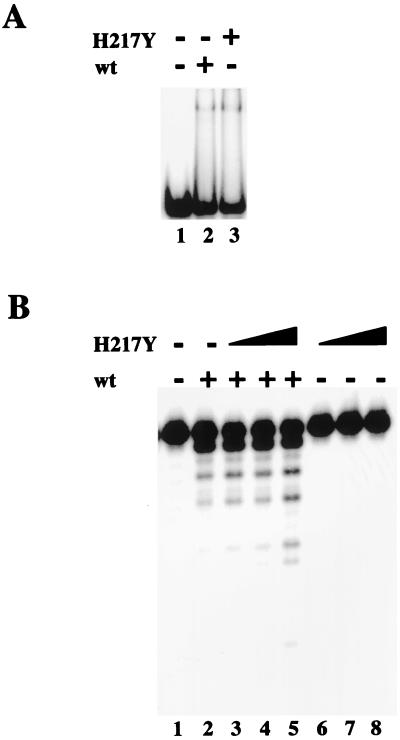
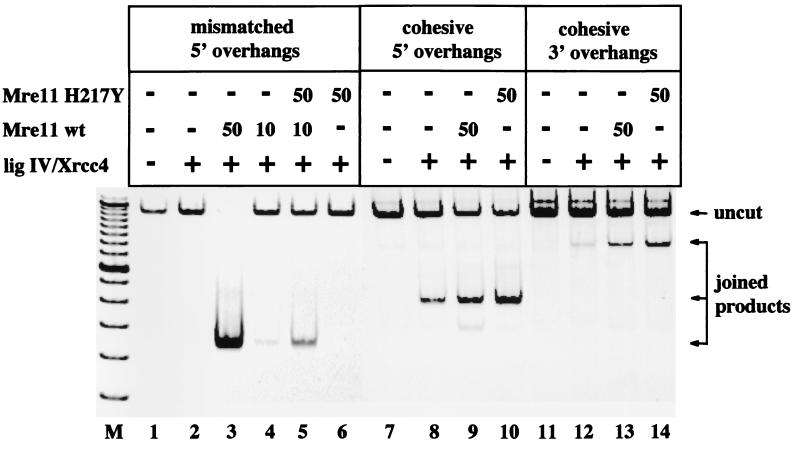
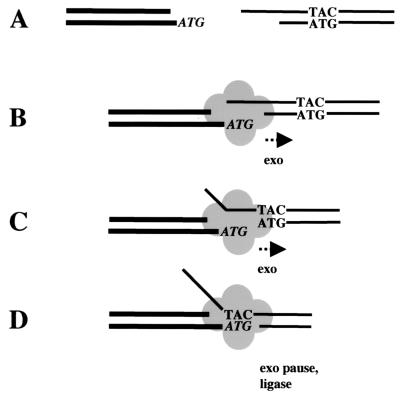
Repair of DNA double-strand breaks in vertebrate cells occurs mainly by an end-joining process that often generates junctions with sequence homologies of a few nucleotides. Mre11 is critical for this mode of repair in budding yeast and has been implicated in the microhomology-based joining. Here, we show that Mre11 exonuclease activity is sensitive to the presence of heterologous DNA, and to the structure and sequence of its ends. Addition of mismatched DNA ends stimulates degradation of DNA by Mre11, whereas cohesive ends strongly inhibit it. Furthermore, if a sequence identity is revealed during the course of degradation, it causes Mre11 nuclease activity to pause, thus stabilizing the junction at a site of microhomology. A nuclease-deficient Mre11 mutant that still binds DNA can also stimulate degradation by wild-type Mre11, suggesting that Mre11-DNA complexes may interact to bridge DNA ends and facilitate DNA joining.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
