

Physiologic determinants of radiation resistance in Deinococcus radiodurans
- PMID: 10831446
- PMCID: PMC110589
- DOI: 10.1128/AEM.66.6.2620-2626.2000
Physiologic determinants of radiation resistance in Deinococcus radiodurans
Abstract
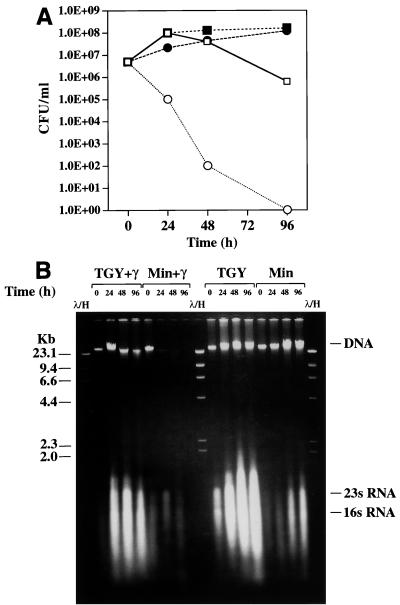
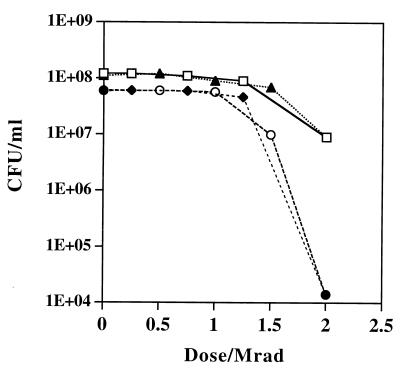
Immense volumes of radioactive wastes, which were generated during nuclear weapons production, were disposed of directly in the ground during the Cold War, a period when national security priorities often surmounted concerns over the environment. The bacterium Deinococcus radiodurans is the most radiation-resistant organism known and is currently being engineered for remediation of the toxic metal and organic components of these environmental wastes. Understanding the biotic potential of D. radiodurans and its global physiological integrity in nutritionally restricted radioactive environments is important in development of this organism for in situ bioremediation. We have previously shown that D. radiodurans can grow on rich medium in the presence of continuous radiation (6,000 rads/h) without lethality. In this study we developed a chemically defined minimal medium that can be used to analyze growth of this organism in the presence and in the absence of continuous radiation; whereas cell growth was not affected in the absence of radiation, cells did not grow and were killed in the presence of continuous radiation. Under nutrient-limiting conditions, DNA repair was found to be limited by the metabolic capabilities of D. radiodurans and not by any nutritionally induced defect in genetic repair. The results of our growth studies and analysis of the complete D. radiodurans genomic sequence support the hypothesis that there are several defects in D. radiodurans global metabolic regulation that limit carbon, nitrogen, and DNA metabolism. We identified key nutritional constituents that restore growth of D. radiodurans in nutritionally limiting radioactive environments.
Figures



Similar articles
-
Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics.Microbiol Mol Biol Rev. 2001 Mar;65(1):44-79. doi: 10.1128/MMBR.65.1.44-79.2001. Microbiol Mol Biol Rev. 2001. PMID: 11238985 Free PMC article. Review.
-
Engineering Deinococcus radiodurans for metal remediation in radioactive mixed waste environments.Nat Biotechnol. 2000 Jan;18(1):85-90. doi: 10.1038/71986. Nat Biotechnol. 2000. PMID: 10625398
-
Why is Deinococcus radiodurans so resistant to ionizing radiation?Trends Microbiol. 1999 Sep;7(9):362-5. doi: 10.1016/s0966-842x(99)01566-8. Trends Microbiol. 1999. PMID: 10470044 Review.
-
Recovery of Deinococcus radiodurans from radiation damage was enhanced under microgravity.Biol Sci Space. 1996 Sep;10(2):97-101. doi: 10.2187/bss.10.97. Biol Sci Space. 1996. PMID: 11785538
-
Repair of ionizing-radiation damage in the radiation resistant bacterium Deinococcus radiodurans.Mutat Res. 1996 May 15;363(1):1-7. doi: 10.1016/0921-8777(95)00014-3. Mutat Res. 1996. PMID: 8632774 Review.
Cited by
-
Responses of Mn2+ speciation in Deinococcus radiodurans and Escherichia coli to γ-radiation by advanced paramagnetic resonance methods.Proc Natl Acad Sci U S A. 2013 Apr 9;110(15):5945-50. doi: 10.1073/pnas.1303376110. Epub 2013 Mar 27. Proc Natl Acad Sci U S A. 2013. PMID: 23536297 Free PMC article.
-
Engineering Deinococcus geothermalis for bioremediation of high-temperature radioactive waste environments.Appl Environ Microbiol. 2003 Aug;69(8):4575-82. doi: 10.1128/AEM.69.8.4575-4582.2003. Appl Environ Microbiol. 2003. PMID: 12902245 Free PMC article.
-
Deinococcus radiodurans Toxin-Antitoxin MazEF-dr Mediates Cell Death in Response to DNA Damage Stress.Front Microbiol. 2017 Jul 26;8:1427. doi: 10.3389/fmicb.2017.01427. eCollection 2017. Front Microbiol. 2017. PMID: 28798741 Free PMC article.
-
Genome analysis and genome-wide proteomics of Thermococcus gammatolerans, the most radioresistant organism known amongst the Archaea.Genome Biol. 2009;10(6):R70. doi: 10.1186/gb-2009-10-6-r70. Epub 2009 Jun 26. Genome Biol. 2009. PMID: 19558674 Free PMC article.
-
cAMP-independent DNA binding of the CRP family protein DdrI from Deinococcus radiodurans.mBio. 2024 Jul 17;15(7):e0114424. doi: 10.1128/mbio.01144-24. Epub 2024 Jun 25. mBio. 2024. PMID: 38916345 Free PMC article.
References
-
- Brim H, McFarlan S C, Fredrickson J K, Minton K W, Zhai M, Wackett L P, Daly M J. Engineering Deinococcus radiodurans for metal remediation in radioactive mixed waste environments. Nature Biotechnol. 2000;18:85–90. - PubMed
-
- Brooks B W, Murray R G E, Johnson J L, Stackebrandt E, Woese C R, Fox G E. Red-pigmented micro cocci: a basis for taxonomy. Int J Sys Bacteriol. 1980;30:627–646.
-
- Cashel M. Detection of (p)ppGpp accumulation patterns in Escherichia coli mutants. Methods Mol Genet. 1994;3:341–356.
-
- Cashel M, Rudd K E. The stringent response in Escherichia coli and Salmonella typhimurium. In: Neidhardt F C, et al., editors. Cellular and molecular biology. Washington, D.C.: American Society for Microbiology; 1996. pp. 1410–1438.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
