

Locus specificity determinants in the multifunctional yeast silencing protein Sir2
- PMID: 10835361
- PMCID: PMC212746
- DOI: 10.1093/emboj/19.11.2641
Locus specificity determinants in the multifunctional yeast silencing protein Sir2
Abstract
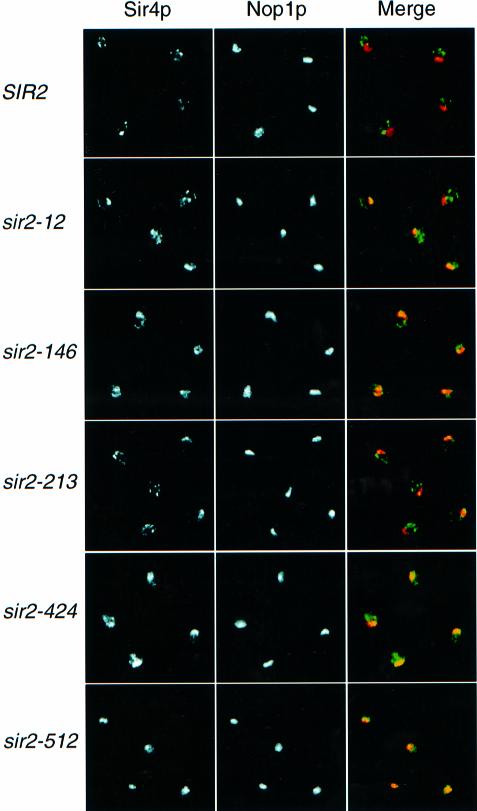
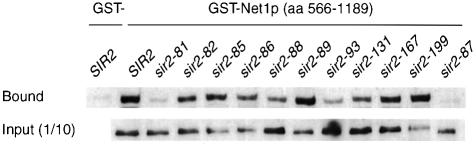
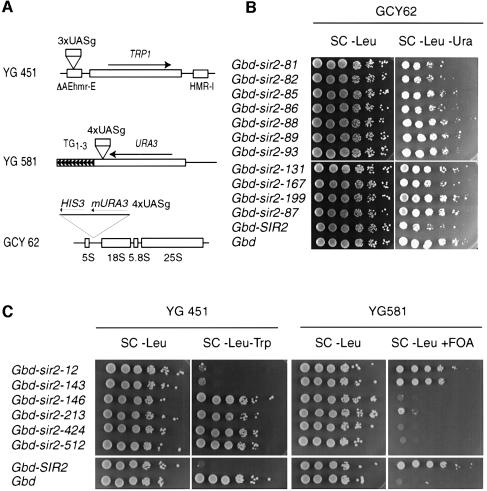
Yeast SIR2, the founding member of a conserved gene family, acts to modulate chromatin structure in three different contexts: silent (HM) mating-type loci, telomeres and rDNA. At HM loci and telomeres, Sir2p forms a complex with Sir3p and Sir4p. However, Sir2p's role in rDNA silencing is Sir3/4 independent, requiring instead an essential nucleolar protein, Net1p. We describe two novel classes of SIR2 mutations specific to either HM/telomere or rDNA silencing. Despite their opposite effects, both classes of mutations cluster in the same two regions of Sir2p, each of which borders on a conserved core domain. A surprising number of these mutations are dominant. Several rDNA silencing mutants display a Sir2p nucleolar localization defect that correlates with reduced Net1p binding. Although the molecular defect in HM/telomere-specific mutants is unclear, they mimic an age-related phenotype where Sir3p and Sir4p relocalize to the nucleolus. Artificial targeting can circumvent the silencing defect in a subset of mutants from both classes. These results define distinct functional domains of Sir2p and provide evidence for additional Sir2p-interacting factors with locus-specific silencing functions.
Figures
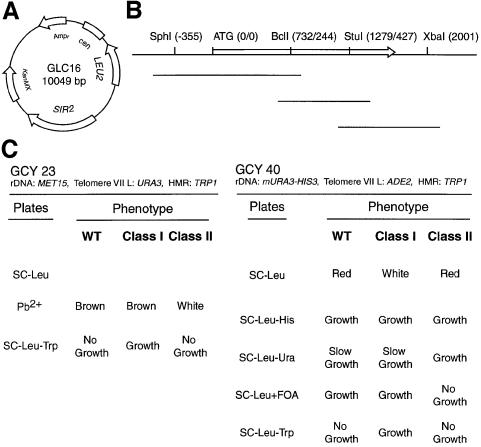


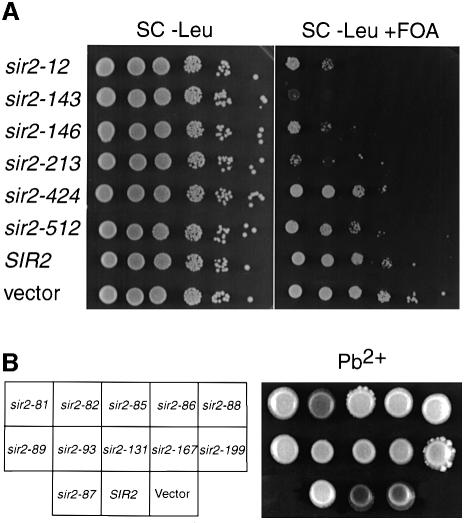
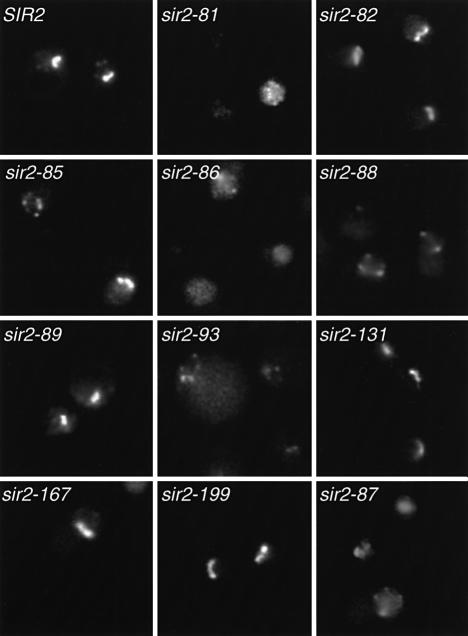
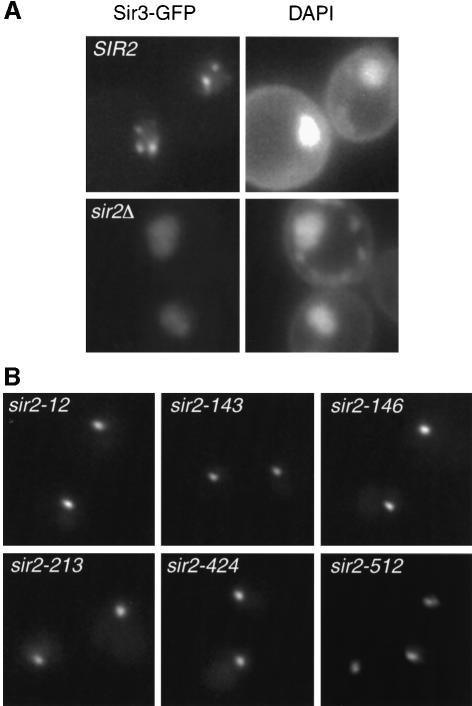
References
-
- Afshar G. and Murnane,J.P. (1999) Characterization of a human gene with sequence homology to Saccharomyces cerevisiae SIR2. Gene, 234, 161–168. - PubMed
-
- Aparicio O.M., Billington,B.L. and Gottschling,D.E. (1991) Modifiers of position effect are shared between telomeric and silent mating-type loci in S.cerevisiae. Cell, 66, 1279–1287. - PubMed
-
- Bartel P.L. and Fields,S. (1995) Analyzing protein–protein interactions using two-hybrid system. Methods Enzymol., 254, 241–263. - PubMed
-
- Brachmann C.B., Sherman,J.M., Devine,S.E., Cameron,E.E., Pillus,L. and Boeke,J.D. (1995) The SIR2 gene family, conserved from bacteria to humans, functions in silencing, cell cycle progression and chromosome stability. Genes Dev., 9, 2888–2902. - PubMed
-
- Brand A.H., Breeden,L., Abraham,J., Sternglanz,R. and Nasmyth,K. (1985) Characterization of a ‘silencer’ in yeast: a DNA sequence with properties opposite to those of a transcriptional enhancer. Cell, 41, 41–48. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
