

gamma-catenin is regulated by the APC tumor suppressor and its oncogenic activity is distinct from that of beta-catenin
- PMID: 10837025
- PMCID: PMC316666
gamma-catenin is regulated by the APC tumor suppressor and its oncogenic activity is distinct from that of beta-catenin
Abstract
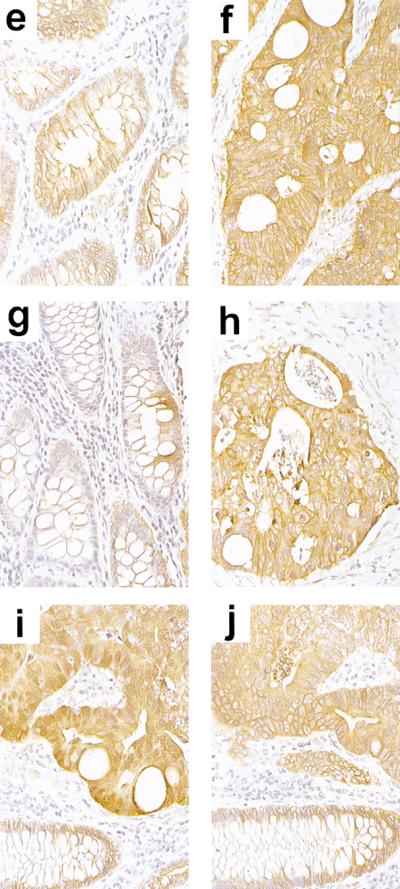
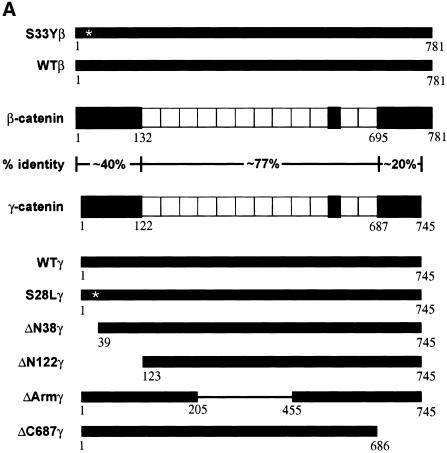
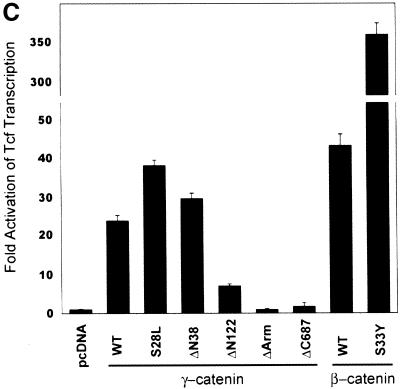
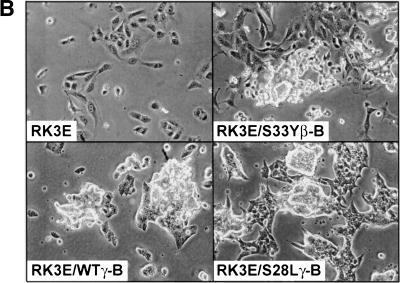
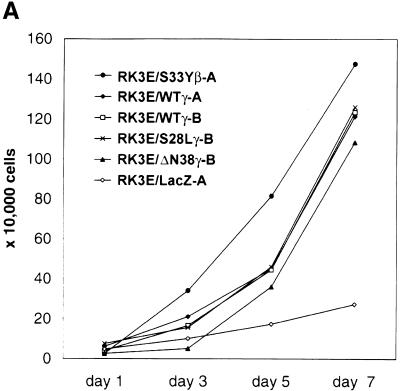
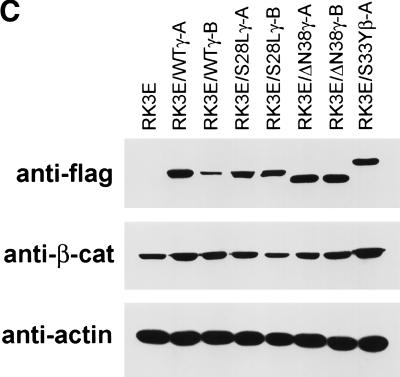
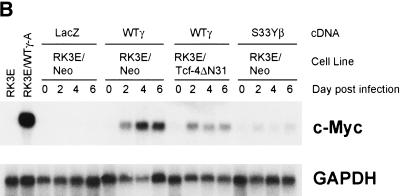
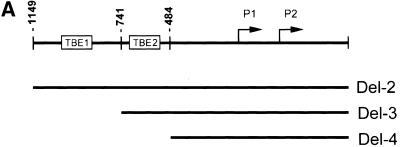
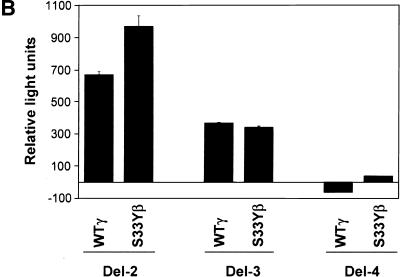
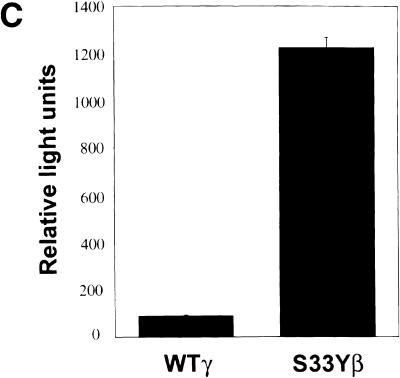
beta-Catenin and gamma-catenin (plakoglobin), vertebrate homologs of Drosophila armadillo, function in cell adhesion and the Wnt signaling pathway. In colon and other cancers, mutations in the APC tumor suppressor protein or beta-catenin's amino terminus stabilize beta-catenin, enhancing its ability to activate transcription of Tcf/Lef target genes. Though beta- and gamma-catenin have analogous structures and functions and like binding to APC, evidence that gamma-catenin has an important role in cancer has been lacking. We report here that APC regulates both beta- and gamma-catenin and gamma-catenin functions as an oncogene. In contrast to beta-catenin, for which only amino-terminal mutated forms transform RK3E epithelial cells, wild-type and several amino-terminal mutated forms of gamma-catenin had similar transforming activity. gamma-Catenin's transforming activity, like beta-catenin's, was dependent on Tcf/Lef function. However, in contrast to beta-catenin, gamma-catenin strongly activated c-Myc expression and c-Myc function was crucial for gamma-catenin transformation. Our findings suggest APC mutations alter regulation of both beta- and gamma-catenin, perhaps explaining why the frequency of APC mutations in colon cancer far exceeds that of beta-catenin mutations. Elevated c-Myc expression in cancers with APC defects may be due to altered regulation of both beta- and gamma-catenin. Furthermore, the data imply beta- and gamma-catenin may have distinct roles in Wnt signaling and cancer via differential effects on downstream target genes.
Figures








Similar articles
-
Beta- and gamma-catenin mutations, but not E-cadherin inactivation, underlie T-cell factor/lymphoid enhancer factor transcriptional deregulation in gastric and pancreatic cancer.Cell Growth Differ. 1999 Jun;10(6):369-76. Cell Growth Differ. 1999. PMID: 10392898
-
Neoplastic transformation of RK3E by mutant beta-catenin requires deregulation of Tcf/Lef transcription but not activation of c-myc expression.Mol Cell Biol. 1999 Aug;19(8):5696-706. doi: 10.1128/MCB.19.8.5696. Mol Cell Biol. 1999. PMID: 10409758 Free PMC article.
-
A comparative evaluation of beta-catenin and plakoglobin signaling activity.Oncogene. 2000 Nov 23;19(50):5720-8. doi: 10.1038/sj.onc.1203921. Oncogene. 2000. PMID: 11126358
-
Catenins, Wnt signaling and cancer.Bioessays. 2000 Nov;22(11):961-5. doi: 10.1002/1521-1878(200011)22:11<961::AID-BIES1>3.0.CO;2-T. Bioessays. 2000. PMID: 11056471 Review.
-
The Yin-Yang of TCF/beta-catenin signaling.Adv Cancer Res. 2000;77:1-24. doi: 10.1016/s0065-230x(08)60783-6. Adv Cancer Res. 2000. PMID: 10549354 Review.
Cited by
-
Human synthetic lethal inference as potential anti-cancer target gene detection.BMC Syst Biol. 2009 Dec 16;3:116. doi: 10.1186/1752-0509-3-116. BMC Syst Biol. 2009. PMID: 20015360 Free PMC article.
-
Restoration of plakoglobin expression in bladder carcinoma cell lines suppresses cell migration and tumorigenic potential.Br J Cancer. 2005 Jun 20;92(12):2153-9. doi: 10.1038/sj.bjc.6602651. Br J Cancer. 2005. PMID: 15942628 Free PMC article.
-
The Prognostic Impact of Protein Expression of E-Cadherin-Catenin Complexes Differs between Rectal and Colon Carcinoma.Gastroenterol Res Pract. 2010;2010:616023. doi: 10.1155/2010/616023. Epub 2010 Aug 16. Gastroenterol Res Pract. 2010. PMID: 20814557 Free PMC article.
-
Desmosomes: a role in cancer?Br J Cancer. 2007 Jun 18;96(12):1783-7. doi: 10.1038/sj.bjc.6603808. Epub 2007 May 22. Br J Cancer. 2007. PMID: 17519903 Free PMC article. Review.
-
Cell-cell contact preserves cell viability via plakoglobin.PLoS One. 2011;6(10):e27064. doi: 10.1371/journal.pone.0027064. Epub 2011 Oct 28. PLoS One. 2011. PMID: 22046445 Free PMC article.
References
-
- Barth AI, Nathke IS, Nelson WJ. Cadherins, catenins and APC protein: Interplay between cytoskeletal complexes and signaling pathways. Curr Opin Cell Biol. 1997;9:683–690. - PubMed
-
- Behrens J. Cadherins and catenins: Role in signal transduction and tumor progression. Cancer Met Rev. 1999;18:15–30. - PubMed
-
- Behrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature. 1996;382:638–642. - PubMed
-
- Ben-Ze'ev A, Geiger B. Differential molecular interactions of β-catenin and plakoglobin in adhesion, signaling and cancer. Curr Opin Cell Biol. 1998;10:629–639. - PubMed
-
- Bienz M. APC: The plot thickens. Curr Opin Genet Dev. 1999;9:595–603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous