

Gross chromosomal rearrangements and genetic exchange between nonhomologous chromosomes following BRCA2 inactivation
- PMID: 10837032
- PMCID: PMC316655
Gross chromosomal rearrangements and genetic exchange between nonhomologous chromosomes following BRCA2 inactivation
Abstract
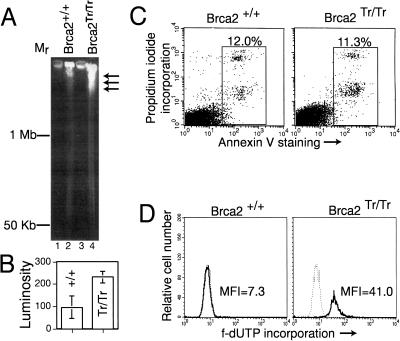
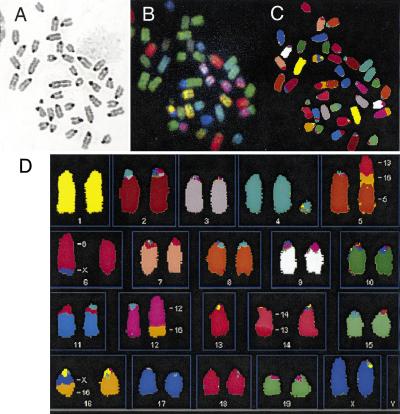
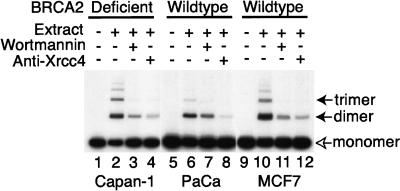
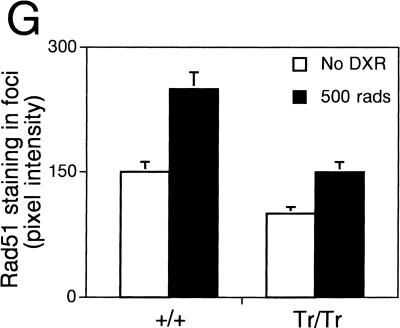
Cancer-causing mutations often arise from gross chromosomal rearrangements (GCRs) such as translocations, which involve genetic exchange between nonhomologous chromosomes. Here we show that murine Brca2 has an essential function in suppressing GCR formation after chromosome breakage. Cells that harbor truncated Brca2 spontaneously incur GCRs and genomic DNA breaks during division. They exhibit hypersensitivity to DNA damage by interstrand cross-linkers, which even at low doses trigger aberrant genetic exchange between nonhomologous chromosomes. Therefore, genetic instability in Brca2-deficient cells results from the mutagenic processing of spontaneous or induced DNA damage into gross chromosomal rearrangements, providing a mechanistic basis for cancer predisposition.
Figures



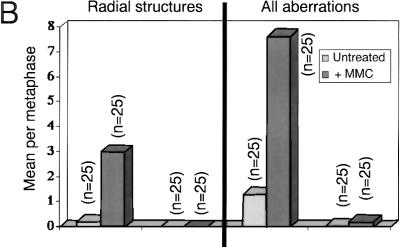
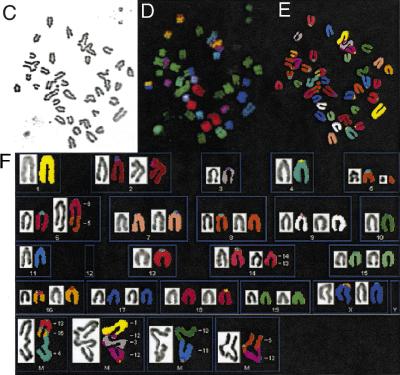
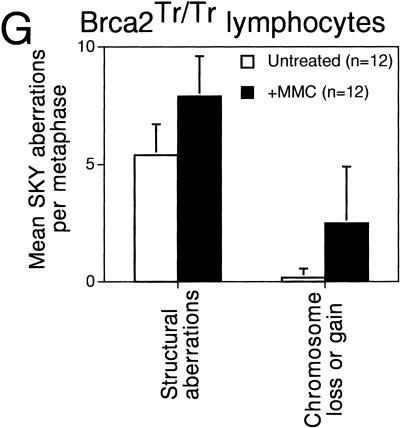
References
-
- Baumann P, Benson FE, West SC. Human Rad51 protein promotes ATP-dependent homologous pairing and strand transfer reactions in vitro. Cell. 1996;87:757–766. - PubMed
-
- Benson FE, Baumann P, West SC. Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature. 1998;391:401–404. - PubMed
-
- Brown DG, Sun XM, Cohen GM. Dexamethasone-induced apoptosis involves cleavage of DNA to large fragments prior to internucleosomal fragmentation. J Biol Chem. 1993;268:3037–3039. - PubMed
-
- Chen C, Kolodner RD. Gross chromosomal rearrangements in S. cerevisiae replication and recombination defective mutants. Nat Genet. 1999;23:81–85. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous