

Receptor selection in B and T lymphocytes
- PMID: 10837051
- PMCID: PMC3822044
- DOI: 10.1146/annurev.immunol.18.1.19
Receptor selection in B and T lymphocytes
Abstract
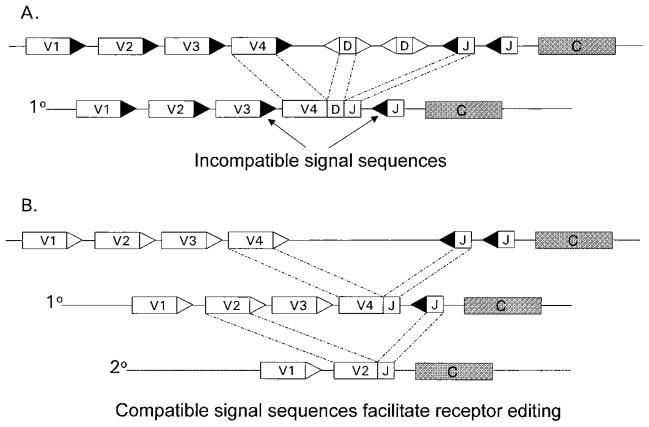
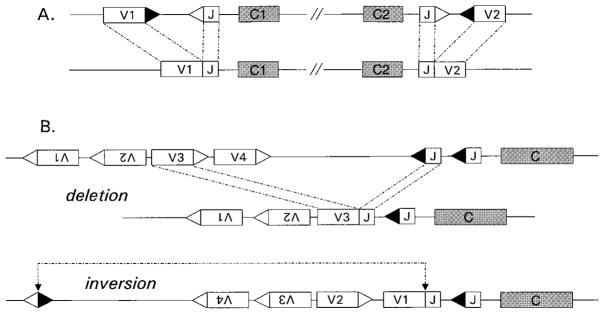
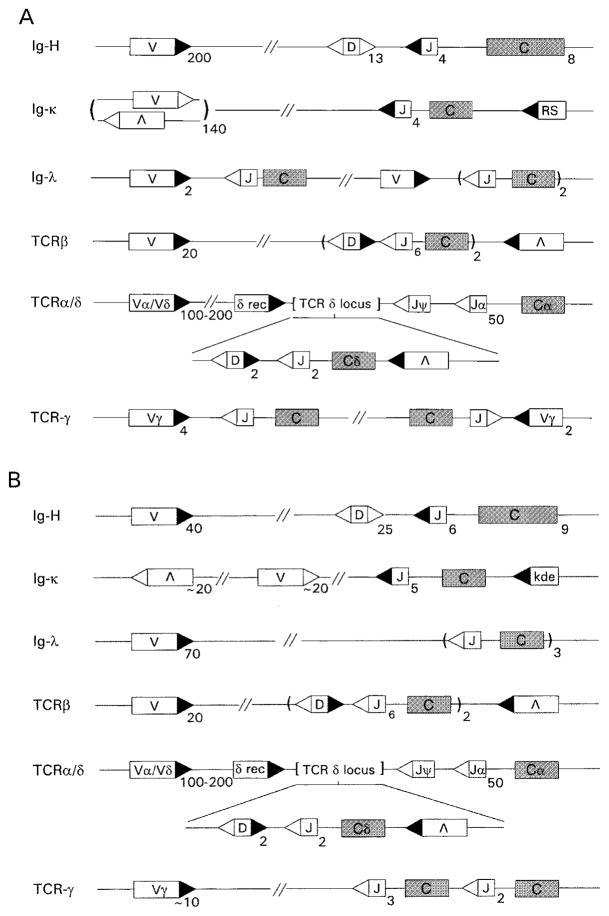
The process of clonal selection is a central feature of the immune system, but immune specificity is also regulated by receptor selection, in which the fate of a lymphocyte's antigen receptor is uncoupled from that of the cell itself. Whereas clonal selection controls cell death or survival in response to antigen receptor signaling, receptor selection regulates the process of V(D)J recombination, which can alter or fix antigen receptor specificity. Receptor selection is carried out in both T and B cells and can occur at different stages of lymphocyte differentiation, in which it plays a key role in allelic exclusion, positive selection, receptor editing, and the diversification of the antigen receptor repertoire. Thus, the immune system takes advantage of its control of V(D)J recombination to modify antigen receptors in such a way that self/non-self discrimination is enhanced. New information about receptor editing in T cells and B-1 B cells is also discussed.
Figures
Similar articles
-
Role of receptor editing and revision in shaping the B and T lymphocyte repertoire.Life Sci. 2001 Jul 27;69(10):1105-13. doi: 10.1016/s0024-3205(01)01219-x. Life Sci. 2001. PMID: 11508343 Review.
-
Immune receptor editing: revise and select.Cell. 1998 Dec 23;95(7):875-8. doi: 10.1016/s0092-8674(00)81711-0. Cell. 1998. PMID: 9875841 Review.
-
Regulation of antigen receptor gene assembly in lymphocytes.Immunol Res. 2001;23(2-3):121-33. doi: 10.1385/IR:23:2-3:121. Immunol Res. 2001. PMID: 11444378 Review.
-
Models for antigen receptor gene rearrangement. I. Biased receptor editing in B cells: implications for allelic exclusion.J Immunol. 1999 Aug 15;163(4):1793-8. J Immunol. 1999. PMID: 10438911
-
Accessibility control of V(D)J recombination.Adv Immunol. 2006;91:45-109. doi: 10.1016/S0065-2776(06)91002-5. Adv Immunol. 2006. PMID: 16938538 Review.
Cited by
-
Antibody repertoire development in fetal and neonatal piglets. XXII. λ Rearrangement precedes κ rearrangement during B-cell lymphogenesis in swine.Immunology. 2012 Oct;137(2):149-59. doi: 10.1111/j.1365-2567.2012.03615.x. Immunology. 2012. PMID: 22724577 Free PMC article.
-
A role for nuclear factor kappa B/rel transcription factors in the regulation of the recombinase activator genes.Immunity. 2005 Apr;22(4):519-31. doi: 10.1016/j.immuni.2005.03.006. Immunity. 2005. PMID: 15845455 Free PMC article.
-
Human autoantibody silencing by immunoglobulin light chains.J Exp Med. 2004 Jul 19;200(2):191-9. doi: 10.1084/jem.20040818. J Exp Med. 2004. PMID: 15263026 Free PMC article.
-
Development and implementation of an automated and highly accurate reporting process for NGS-based clonality testing.Oncotarget. 2023 May 12;14:450-461. doi: 10.18632/oncotarget.28429. Oncotarget. 2023. PMID: 37171376 Free PMC article.
-
Phospholipase Cgamma2 contributes to light-chain gene activation and receptor editing.Mol Cell Biol. 2007 Sep;27(17):5957-67. doi: 10.1128/MCB.02273-06. Epub 2007 Jun 25. Mol Cell Biol. 2007. PMID: 17591700 Free PMC article.
References
-
- Radic MZ, Zouali M. Receptor editing, immune diversification, and self-tolerance. Immunity. 1996;5:505–11. - PubMed
-
- Hertz M, Nemazee D. Receptor editing and commitment in B lymphocytes. Curr Opin Immunol. 1998;10:208–13. - PubMed
-
- Nussenzweig MC. Immune receptor editing: revise and select. Cell. 1998;95:875–78. - PubMed
-
- Ohmori H, Hikida M. Expression and function of recombination activating genes in mature B cells. Crit Rev Immunol. 1998;18:221–35. - PubMed
-
- Kofler R, Geley S, Kofler H, Helmberg A. Mouse variable-region gene families: complexity, polymorphism and use in non-autoimmune responses. Immunol Rev. 1992;128:5–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
