

Mechanisms of enveloped virus entry into animal cells
- PMID: 10837671
- PMCID: PMC7125647
- DOI: 10.1016/s0169-409x(98)00002-7
Mechanisms of enveloped virus entry into animal cells
Abstract
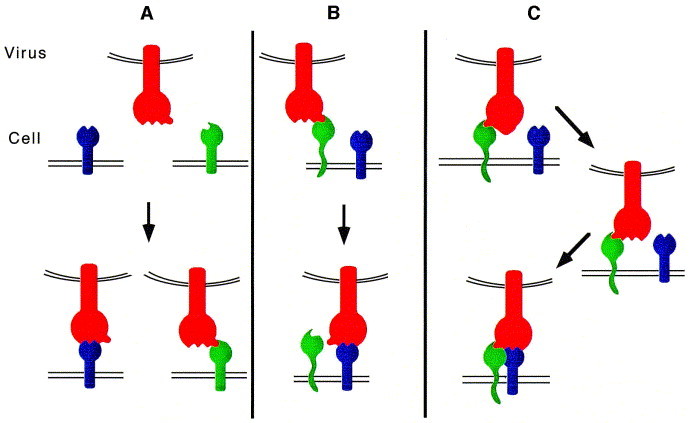
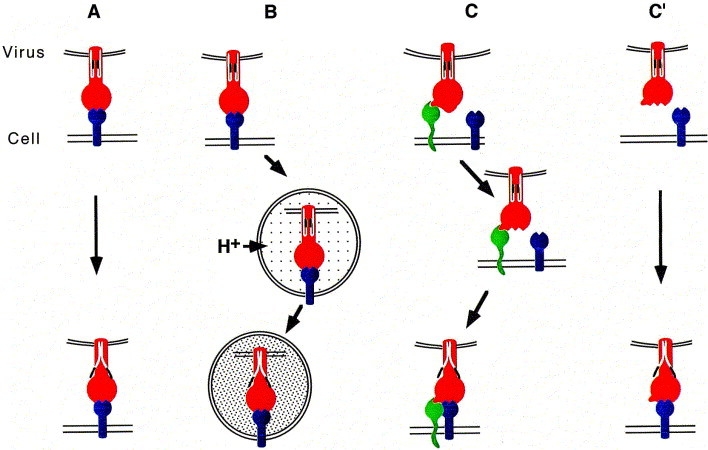
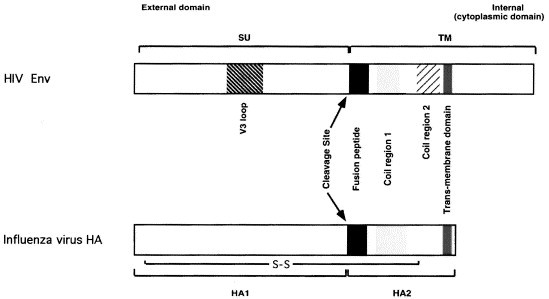
The ability of viruses to transfer macromolecules between cells makes them attractive starting points for the design of biological delivery vehicles. Virus-based vectors and sub-viral systems are already finding biotechnological and medical applications for gene, peptide, vaccine and drug delivery. Progress has been made in understanding the cellular and molecular mechanisms underlying virus entry, particularly in identifying virus receptors. However, receptor binding is only a first step and we now have to understand how these molecules facilitate entry, how enveloped viruses fuse with cells or non-enveloped viruses penetrate the cell membrane, and what happens following penetration. Only through these detailed analyses will the full potential of viruses as vectors and delivery vehicles be realised. Here we discuss aspects of the entry mechanisms for several well-characterised viral systems. We do not attempt to provide a fully comprehensive review of virus entry but focus primarily on enveloped viruses.
Figures
References
-
- Adams L.D, Tomasselli A.G, Robbins P, Moss B, Heinrikson R.L. HIV-1 protease cleaves actin during acute infection of human T-lymphocytes. AIDS Res. Hum. Retroviruses. 1992;8:291–295. - PubMed
-
- Albritton L.M, Tseng L, Scadden D, Cunningham J.M. A putative murine retrovirus receptor gene encodes a multiple membrane spanning protein and confers susceptibility to virus infection. Cell. 1989;57:659–666. - PubMed
-
- Andersen K.B, Skov H. Retrovirus-induced cell fusion is enhanced by protease treatment. J. Gen. Virol. 1989;70:1921–1927. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
