

Investigation of routes and funnels in protein folding by free energy functional methods
- PMID: 10841554
- PMCID: PMC18640
- DOI: 10.1073/pnas.97.12.6509
Investigation of routes and funnels in protein folding by free energy functional methods
Abstract
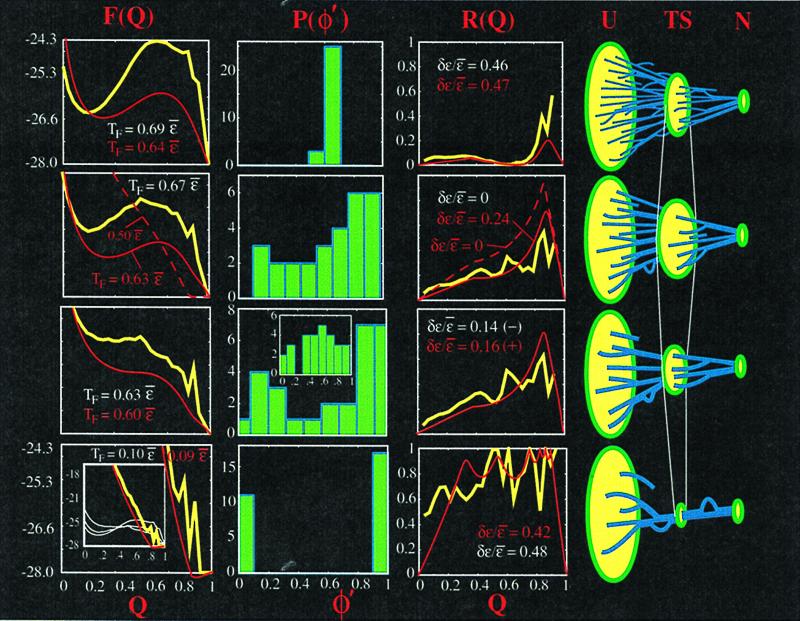
We use a free energy functional theory to elucidate general properties of heterogeneously ordering, fast folding proteins, and we test our conclusions with lattice simulations. We find that both structural and energetic heterogeneity can lower the free energy barrier to folding. Correlating stronger contact energies with entropically likely contacts of a given native structure lowers the barrier, and anticorrelating the energies has the reverse effect. Designing in relatively mild energetic heterogeneity can eliminate the barrier completely at the transition temperature. Sequences with native energies tuned to fold uniformly, as well as sequences tuned to fold reliably by a single or a few routes, are rare. Sequences with weak native energetic heterogeneity are more common; their folding kinetics is more strongly determined by properties of the native structure. Sequences with different distributions of stability throughout the protein may still be good folders to the same structure. A measure of folding route narrowness is introduced that correlates with rate and that can give information about the intrinsic biases in ordering arising from native topology. This theoretical framework allows us to investigate systematically the coupled effects of energy and topology in protein folding and to interpret recent experiments that investigate these effects.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
